Bir gen bir ferment zamonaviy talqini. Bitta gen - bitta ferment nazariyasi
4.2.1. "Bir gen, bitta ferment" gipotezasi
Birinchi tadqiqot. Garrod 1902 yilda alkaptonuriyadagi irsiy nuqson organizmning gomogentis kislotasini parchalay olmasligi bilan bog'liqligini ta'kidlaganidan so'ng, bu buzuqlikning o'ziga xos mexanizmini tushuntirish muhim edi. O'sha paytda metabolik reaktsiyalar fermentlar tomonidan katalizlanganligi ma'lum bo'lganligi sababli, alkaptonuriyaga olib kelgan ba'zi fermentlarning buzilishi deb taxmin qilish mumkin edi. Bunday gipotezani Driesch (1896 yilda) muhokama qilgan. Buni Xelden (1920, qarang) va Garrod (1923) ham ifodalagan. Muhim bosqichlar Biokimyoviy genetikaning rivojlanishi Kuchn va Butenandtning tegirmon kuyalarida ko'z rangini o'rganish bo'yicha ishlaridan boshlangan. Efestiya kuhniella va Beadle va Ephrussi tomonidan shunga o'xshash tadqiqotlar Drosophila(1936). Ushbu kashshof ishlarda gen ta'sirining mexanizmlarini yoritish uchun avval genetik usullar bilan o'rganilgan hasharotlar mutantlari tanlangan. Biroq, bu yondashuv muvaffaqiyatga olib kelmadi. Muammo juda murakkab bo'lib chiqdi va uni hal qilish kerak edi:
1) eksperimental o'rganish uchun qulay bo'lgan oddiy model organizmni tanlash;
2) genetik jihatdan aniqlangan belgilarning biokimyoviy asoslarini emas, balki biokimyoviy belgilarning genetik asoslarini izlash. 1941 yilda Bidl va Tatum ishida ikkala shart ham bajarilgan (shuningdek qarang: Beadle, 1945).
Beadle va Tatumning modeli. Ushbu tadqiqotchilarning maqolasi shunday boshlangan:
“Fiziologik genetika nuqtai nazaridan organizmning rivojlanishi va faoliyati qandaydir tarzda genlar tomonidan boshqariladigan murakkab kimyoviy reaksiyalar tizimiga aylanishi mumkin. Bu genlar... yoki o'zlari ferment vazifasini bajaradi yoki ularning o'ziga xosligini aniqlaydi, deb taxmin qilish juda mantiqiy. Ma'lumki, genetik-fiziologlar odatda allaqachon ma'lum bo'lgan irsiy xususiyatlarning fiziologik va biokimyoviy asoslarini tekshirishga harakat qilishadi. Ushbu yondashuv juda ko'p bioni yaratishga imkon berdi kimyoviy reaksiyalar maxsus genlar tomonidan boshqariladi. Bunday tadqiqotlar fermentlar va genlar bir xil tartibdagi o'ziga xoslikka ega ekanligini ko'rsatdi. Biroq, bu yondashuvning imkoniyatlari cheklangan. Eng jiddiy cheklov shundaki, bu holda tadqiqotchilar halokatli ta'sirga ega bo'lmagan va shuning uchun organizmning hayoti uchun juda muhim bo'lmagan reaktsiyalar bilan bog'liq bo'lgan irsiy belgilarga e'tibor berishadi. Ikkinchi qiyinchilik... muammoga anʼanaviy yondashuv tashqi koʻrinishdagi belgilardan foydalanishni nazarda tutadi. Ularning aksariyati biokimyoviy reaktsiya tizimlariga asoslangan morfologik o'zgarishlar shu qadar murakkabki, ularni tahlil qilish juda qiyin.
Shu kabi mulohazalar bizni quyidagi xulosaga olib keldi. Rivojlanish va metabolizmni belgilaydigan biokimyoviy reaktsiyalarni genetik nazorat qilishning umumiy muammosini o'rganish quyidagi usullardan foydalangan holda amalga oshirilishi kerak. umumiy qabul qilinganga qarama-qarshi protsedura: Ma'lum bo'lgan irsiy xususiyatlarning kimyoviy asoslarini aniqlashga harakat qilish o'rniga, uni aniqlash kerak genlar ma'lum biokimyoviy reaktsiyalarni nazorat qiladimi va qanday. Askomitsetlarga tegishli bo'lgan neyrospora bunday yondashuvni amalga oshirishga imkon beradigan xususiyatlarga ega va ayni paytda genetik tadqiqotlar uchun qulay ob'ekt bo'lib xizmat qiladi. Shuning uchun bizning dasturimiz ushbu organizmdan foydalanishga asoslangan. Biz rentgen nurlari ma'lum kimyoviy reaktsiyalarni boshqaradigan genlarda mutatsiyaga olib keladi deb taxmin qildik. Aytaylik, ma'lum bir muhitda omon qolish uchun organizm qandaydir kimyoviy reaktsiyani amalga oshirishi kerak, unda bunday qobiliyatga ega bo'lmagan mutant bu sharoitda yaroqsiz bo'ladi. Biroq, agar genetik jihatdan bloklangan reaktsiyaning hayotiy mahsuloti qo'shilgan muhitda o'stirilsa, uni saqlab qolish va o'rganish mumkin.
4 Genlarning harakati 9
Guruch. 4.1. Neyrosporaning biokimyoviy mutantlarini aniqlash bo'yicha eksperiment sxemasi To'liq muhitda rentgen nurlari yoki ultrabinafsha nurlanish bilan qo'zg'atilgan mutatsiyalar qo'ziqorin o'sishini buzmaydi. Biroq, mutant minimal muhitda o'smaydi. Vitaminlar minimal muhitga qo'shilsa, o'sish qobiliyati tiklanadi.Aminokislotalar qo'shilsa, o'sish bo'lmaydi.Bu ma'lumotlarga asoslanib, mutatsiya vitamin almashinuvini boshqaruvchi genda sodir bo'lgan deb taxmin qilish mumkin. Keyingi qadam normal funktsiyani tiklay oladigan vitaminni aniqlashdir Vitamin biosintezi reaktsiyalari orasida genetik blok mavjud.Keyinchalik, Beadle va Tatum eksperimental dizaynni tasvirlaydi (4.1-rasm). To'liq muhitda agar, noorganik tuzlar, solod ekstrakti, xamirturush ekstrakti va glyukoza mavjud edi. Minimal muhitda faqat agar, tuzlar, biotin va uglerod manbai mavjud edi. To'liq muhitda o'sgan va minimal muhitda o'smagan mutantlar eng batafsil o'rganildi. Mutantlarning har birida sintezi buzilgan birikmani aniqlash uchun minimal agarga to'liq muhitning alohida komponentlari qo'shildi.
Shu tarzda, ma'lum o'sish omillarini: piridoksin, tiamin va para-aminobenzoy kislotasini sintez qila olmaydigan shtammlar ajratildi. Bu nuqsonlar ma’lum lokuslardagi mutatsiyalar natijasida yuzaga kelganligi isbotlangan. Ish neyrospora, bakteriyalar va xamirturushlar bo'yicha ko'plab tadqiqotlar uchun asos yaratdi, ularda metabolizmning individual bosqichlari va o'ziga xos ferment buzilishlari uchun javob beradigan "genetik bloklar" ning muvofiqligi aniqlandi. Ushbu yondashuv tezda tadqiqotchilarga metabolik yo'llarni ochishga imkon beruvchi vositaga aylandi.
"Bir gen - bitta ferment" gipotezasi kuchli eksperimental tasdiqni oldi. Keyingi o'n yilliklar ishi ko'rsatganidek, bu hayratlanarli darajada samarali bo'ldi. Buzuq fermentlar va ularning normal variantlarini tahlil qilish tez orada ferment funktsiyasining o'zgarishiga olib keladigan genetik kasalliklar sinfini aniqlash imkonini berdi, garchi oqsilning o'zi hali ham aniqlansa va immunologik xususiyatlarini saqlab qoldi. Boshqa hollarda, ferment faolligi uchun optimal harorat o'zgaradi. Ba'zi variantlarni umumiy tartibga solish mexanizmiga ta'sir qiluvchi mutatsiya va natijada butun fermentlar guruhining faolligini o'zgartirishi bilan izohlash mumkin. Shunga o'xshash tadqiqotlar operon tushunchasini o'z ichiga olgan bakteriyalarda gen faolligini tartibga solish kontseptsiyasini yaratishga olib keldi.
10 4. Genlarning harakati
Odamlarda fermentativ buzilishlarning birinchi misollari. Enzimatik buzilish namoyon bo'lishi mumkin bo'lgan birinchi irsiy odam kasalligi retsessiv irsiy usul bilan methemoglobinemiya edi (Gibson va Xarrison, 1947; Gibson, 1948) (25080). Bunday holda, shikastlangan ferment NADH - qaram methemoglobin reduktaza hisoblanadi. Metabolik nuqsonlar bilan bog'liq bo'lgan inson kasalliklari guruhini tizimli ravishda o'rganishga birinchi urinish 1951 yilda qilingan. Glikogenni saqlash kasalligini o'rganishda Koreys Gierke kasalligi (23220) deb tashxis qo'yilgan patologik holatning o'nta holatidan sakkiztasida jigar glikogen tuzilishi normal ekanligini va ikkita holatda aniq anormal ekanligini ko'rsatdi. Bundan tashqari, ortiqcha to'plangan jigar glikogenini to'g'ridan-to'g'ri shakarga aylantira olmasligi aniq edi, chunki bemorlar gipoglikemiyaga moyil edi. Jigarda glyukoza hosil qilish uchun glikogenni parchalash uchun ko'plab fermentlar kerak. Ulardan ikkitasi, amilo-1,6-glyukozidaza va glyukoza 6-fosfataza ferment tizimining mumkin bo'lgan nuqsonli elementlari sifatida o'rganish uchun tanlangan. Glyukoza-6fosfatdan fosfatning chiqarilishi jigar homogenatlarida turli pH qiymatlarida o'lchandi. Natijalar rasmda keltirilgan. 4.2. Oddiy jigarda yuqori faollik pH 6-7 da tegmaslik bilan topilgan. Sirozda jiddiy jigar disfunktsiyasi faollikning biroz pasayishi bilan bog'liq. Boshqa tomondan, o'limga olib keladigan Gierke kasalligi bo'lsa, ferment faolligini umuman aniqlab bo'lmadi; xuddi shunday natija ikkinchi shunga o'xshash bemorni tekshirganda olingan. Kamroq alomatlari bo'lgan ikkita bemorda faollik sezilarli darajada pasaygan.
O'limga olib keladigan Gierke kasalligining bu holatlarida glyukoza-6-fosfataza etishmovchiligi mavjud degan xulosaga keldi. Biroq, ko'pchilik engil holatlarda, bu fermentning faolligi jigar sirroziga qaraganda past emas edi va faqat ikkita bemorda u biroz pastroq edi (4.2-rasm).
Korilarning fikriga ko'ra, mushak to'qimalarida glikogenning g'ayritabiiy to'planishi glyukoza-6-fosfataza etishmovchiligi bilan bog'liq bo'lishi mumkin emas, chunki bu ferment mushaklarda yo'q va normaldir. Mushak glikogenozining mumkin bo'lgan izohi sifatida ular amilo-1,6-glyukozidaza faolligining buzilishini taklif qilishdi. Tez orada bu bashorat tasdiqlandi: Forbes yurak va skelet mushaklari bilan bog'liq bo'lgan glikogenni saqlash kasalligining klinik ahamiyatli holatlaridan birida bunday nuqsonni aniqladi. Endi biz
4. Genlarning harakati 11
Glikogenni saqlash kasalligida ko'plab fermentativ nuqsonlar ma'lum.
Ushbu kasallikning turli shakllari namoyon bo'lish darajasida bir oz farq qilsa-da, klinik jihatdan ularning umumiy jihatlari juda ko'p. Bitta istisno bilan ularning barchasi avtosomal retsessiv tarzda meros bo'lib o'tadi. Agar fermentativ nuqsonlar aniqlanmagan bo'lsa, glikogenni saqlash patologiyasi og'irlik darajasi, simptomatik tafsilotlar va o'lim vaqti bo'yicha xarakterli oila ichidagi korrelyatsiyaga ega bo'lgan kasallik sifatida qaraladi. Shunday qilib, bizning oldimizda faqat fenotipni o'rganish (3.3.5-bo'lim) asosida taxmin qilinishi mumkin bo'lgan genetik heterojenlik biokimyoviy darajadagi tahlil bilan tasdiqlangan misol mavjud: tadqiqot fermentativ faollik maxsus genlarni aniqlash imkonini berdi.
Keyingi yillarda fermentativ nuqsonlar sohasidagi tadqiqotlar sur'ati oshdi va MakKusik o'zining "Odamda Mendel merosi" (1983) kitobining oltinchi nashrida tasvirlagan 588 aniqlangan retsessiv autosomal genlar uchun o'ziga xos fermentativ nuqsonlar aniqlandi. 170 ta holat. Bizning bu sohadagi yutuqlarimiz molekulyar genetika tushunchalari va usullarini ishlab chiqish bilan bevosita bog'liq.
Odamlarda fermentativ buzilishlarni o'rganishning ayrim bosqichlari. Biz ushbu davom etayotgan jarayonning faqat eng muhim bosqichlarini taqdim etamiz: 1934 yilda Fölling fenilketonuriyani aniqladi.
1941 yil Beadle va Tatum "bitta gen, bitta ferment" gipotezasini shakllantirdilar 1948 yil Gibson inson kasalliklarida (retsessiv methemoglobinemiya) birinchi fermentativ buzilish holatini tasvirlab berdi.
1952 yil Korilar Gierke kasalligida glyukoza-6-fosfataza etishmovchiligini aniqladilar.
1953 yil Jervis fenilketonuriyada fenilalanin gidroksilaza yo'qligini ko'rsatdi. Bikel, kam fenilalaninli dietadan foydalangan holda fermentativ buzilishni yumshatish uchun birinchi urinish haqida xabar berdi
1955-yil Smithies kraxmalli gel elektroforez texnikasini ishlab chiqdi
1956 yil Karson va boshqalar qo'zg'atilgan gemolitik anemiya holatida glyukoza-6-fosfat dehidrogenaza (G6PD) etishmovchiligini aniqladilar.
1957 yil Kalkar va boshqalar galaktozemiyadagi ferment etishmovchiligini tavsiflab, odamlar va bakteriyalar fermentativ faollikda bir xil nuqsonlarga ega ekanligini ko'rsatdilar.
1961 yil Krut va Vaynberg ekilgan fibroblastlarda in vitro galaktozemiyada ferment nuqsonini ko'rsatdilar.
1967 yil Seegmiller va boshqalar Lesch-Nyhan sindromida gipoksantin-guanin fosforiboziltransferaza (HPRT) nuqsonini aniqladilar.
1968 Kliver kseroderma pigmentosumda eksizyonni tiklashning buzilishini tasvirlab berdi
1970 yil Noyfeld mukopolisaxaridozlarda fermentativ nuqsonlarni aniqladi, bu esa mukopolisaxaridlarning parchalanish yo'llarini aniqlash imkonini berdi.
1974 yil Braun va Goldshteyn oilaviy giperkolesterolemiyada gidroksimetilglutarilKoA-reduktazaning genetik jihatdan aniqlangan ortiqcha ishlab chiqarilishi ushbu fermentning (HMG) faolligini modulyatsiya qiluvchi membranada lokalizatsiya qilingan past zichlikdagi lipoprotein retseptorlaridagi nuqson tufayli ekanligini isbotladi.
1977 Sly va boshqalar mannoz-6-fosfat (lizosomal fermentlarning tarkibiy qismi sifatida) fibroblast retseptorlari tomonidan tan olinishini ko'rsatdi. Genetik qayta ishlash nuqsoni lizosomal fermentlarning bog'lanishiga to'sqinlik qiladi, buning natijasida ularning sitoplazmaga chiqishi va keyinchalik plazmaga sekretsiyasi buziladi (I-hujayra kasalligi)
12 4. Genlarning harakati
1980 Psevdogipoparatiroidizmda retseptor va siklaza bog'lanishini ta'minlaydigan oqsildagi nuqson aniqlandi.
Har bir tirik hujayrada ko'plab kimyoviy reaktsiyalar sodir bo'ladi. Fermentlar maxsus va o'ta muhim funktsiyalarga ega bo'lgan oqsillardir. Ular biokatalizatorlar deb ataladi. Tanadagi asosiy narsa biokimyoviy reaktsiyalarni tezlashtirishdir. O'zaro ta'siri bu molekulalar tomonidan katalizlanadigan boshlang'ich reagentlar substratlar, yakuniy birikmalar esa mahsulotlar deb ataladi.
Tabiatda ferment oqsillari faqat tirik tizimlarda ishlaydi. Ammo zamonaviy biotexnologiyada klinik diagnostika, farmatsevtika va tibbiyot, tozalangan fermentlar yoki ularning komplekslari, shuningdek tizimning ishlashi va tadqiqotchi uchun ma'lumotlarni vizualizatsiya qilish uchun zarur bo'lgan qo'shimcha komponentlar qo'llaniladi.
Fermentlarning biologik ahamiyati va xossalari
Bu molekulalarsiz tirik organizm ishlay olmaydi. Barcha hayotiy jarayonlar fermentlar tufayli muammosiz ishlaydi. Organizmdagi ferment oqsillarining asosiy vazifasi metabolizmni tartibga solishdir. Ularsiz normal metabolizm mumkin emas. Molekulalar faoliyatini tartibga solish aktivatorlar (induktorlar) yoki inhibitorlar ta'sirida sodir bo'ladi. Boshqaruv ishlaydi turli darajalar oqsil sintezi. Shuningdek, u tayyor molekulada "ishlaydi".
Ferment oqsillarining asosiy xossalari ma'lum bir substratga xoslikdir. Va shunga ko'ra, faqat bitta yoki kamroq tez-tez bir qator reaktsiyalarni katalizlash qobiliyati. Odatda bunday jarayonlar teskari bo'ladi. Bitta ferment ikkala funktsiya uchun javobgardir. Lekin bu hammasi emas.

Ferment oqsillarining roli juda muhimdir. Ularsiz biokimyoviy reaktsiyalar sodir bo'lmaydi. Fermentlarning ta'siri tufayli reagentlar sezilarli energiya sarfisiz faollashuv to'sig'ini engib o'tishlari mumkin bo'ladi. Tananing 100 ° C dan yuqori haroratni qizdirishi yoki agressiv komponentlardan foydalanish mumkin emas, masalan. kimyoviy laboratoriya. Ferment oqsili substrat bilan bog'lanadi. Bog'langan holatda modifikatsiya ikkinchisining keyingi chiqarilishi bilan sodir bo'ladi. Kimyoviy sintezda ishlatiladigan barcha katalizatorlar aynan shunday ishlaydi.
Oqsil-ferment molekulasining tashkiliy darajalari qanday?
Odatda bu molekulalar uchinchi (globula) yoki to'rtlamchi (bir nechta bog'langan globulalar) mavjud. oqsil tuzilishi. Ular birinchi navbatda chiziqli shaklda sintezlanadi. Va keyin ular kerakli tuzilishga tushadilar. Faoliyatni ta'minlash uchun biokatalizator ma'lum bir tuzilishga muhtoj.

Fermentlar, boshqa oqsillar kabi, issiqlik, ekstremal pH qiymatlari va agressiv kimyoviy birikmalar bilan yo'q qilinadi.
Fermentlarning qo'shimcha xossalari
Ular orasida tarkibiy qismlarning quyidagi xususiyatlari mavjud:
- Stereospesifiklik - faqat bitta mahsulotning shakllanishi.
- Regioselektivlik - bo'shliq kimyoviy bog'lanish yoki guruhni faqat bitta pozitsiyada o'zgartirish.
- Kimyoselektivlik - faqat bitta reaktsiyaning katalizi.
Ishning xususiyatlari
Darajasi har xil. Lekin har qanday ferment hamisha tuzilish jihatidan oʻxshash boʻlgan maʼlum bir substrat yoki birikmalar guruhiga nisbatan faoldir. Protein bo'lmagan katalizatorlar bunday xususiyatga ega emas. O'ziga xoslik 10−10 mol/L ga yetishi mumkin bo'lgan bog'lanish konstantasi (mol/L) bilan o'lchanadi. Faol fermentning ishi tezdir. Bitta molekula soniyada minglab millionlab operatsiyalarni katalizlaydi. Biokimyoviy reaksiyalarning tezlanish darajasi an'anaviy katalizatorlarga qaraganda sezilarli darajada (1000-100000 marta) yuqori.
Fermentlarning ta'siri bir necha mexanizmlarga asoslanadi. Eng oddiy o'zaro ta'sir bir substrat molekulasi bilan sodir bo'ladi va undan keyin mahsulot hosil bo'ladi. Aksariyat fermentlar reaktsiyaga kiradigan 2-3 xil molekulalarni bog'lash qobiliyatiga ega. Masalan, guruh yoki atomni bir birikmadan ikkinchisiga o'tkazish yoki "ping-pong" tamoyili yordamida ikki marta almashtirish. Ushbu reaksiyalarda odatda bitta substrat birlashtiriladi, ikkinchisi esa funktsional guruh orqali ferment bilan bog'lanadi.
Tadqiqot quyidagi usullar yordamida amalga oshiriladi:
- Oraliq va yakuniy mahsulotlarning ta'riflari.
- Substrat bilan bog'liq struktura va funktsional guruhlarning geometriyasini o'rganish va yuqori ta'minlash
- Ferment genlarining mutatsiyalari va uning sintezi va faolligidagi o'zgarishlarni aniqlash.

Faol va majburiy sayt
Substrat molekulasi o'lchamiga ko'ra ferment oqsilidan ancha kichikdir. Shuning uchun bog'lanish biokatalizatorning oz sonli funktsional guruhlari tufayli sodir bo'ladi. Ular aminokislotalarning ma'lum bir to'plamidan iborat faol markazni hosil qiladi. Strukturada oqsil bo'lmagan tabiatning protez guruhi mavjud bo'lib, u ham faol markazning bir qismi bo'lishi mumkin.
Fermentlarning alohida guruhini ajratish kerak. Ularning molekulasida molekula bilan doimo bog'lanib, undan ajralib chiqadigan koenzim mavjud. To'liq hosil bo'lgan ferment oqsili goloferment, kofaktor olib tashlanganida esa apoenzim deyiladi. Vitaminlar, metallar, azotli asoslarning hosilalari (NAD - nikotinamid adenin dinukleotid, FAD - flavin adenin dinukleotid, FMN - flavin mononukleotid) ko'pincha koferment sifatida ishlaydi.

Bog'lanish joyi substratning o'ziga xosligini ta'minlaydi. Uning hisobidan barqaror substrat-ferment kompleksi hosil bo'ladi. Globulaning tuzilishi sirtda ma'lum o'lchamdagi uyaga (yoriq yoki depressiya) ega bo'lib, substratning bog'lanishini ta'minlaydigan tarzda qurilgan. Bu zona odatda faol markaz yaqinida joylashgan. Alohida fermentlar kofaktorlar yoki metall ionlari bilan bog'lanish joylariga ega.
Xulosa
Enzim oqsili organizmda muhim rol o'ynaydi. Bunday moddalar kimyoviy reaktsiyalarni katalizlaydi va metabolik jarayon - metabolizm uchun javobgardir. Har qanday tirik hujayrada yuzlab biokimyoviy jarayonlar doimo sodir bo'ladi, shu jumladan kamaytirish reaktsiyalari, birikmalarning parchalanishi va sintezi. Moddalar doimo katta miqdorda energiya chiqishi bilan oksidlanadi. U, o'z navbatida, uglevodlar, oqsillar, yog'lar va ularning komplekslarini shakllantirishga sarflanadi. Parchalanish mahsulotlari zarur organik birikmalarni sintez qilish uchun tuzilmaviy elementlardir.
» , » Bitta gen, bitta ferment
Bitta gen, bitta ferment
92
Nashr qilingan sana: 2018 yil 24 iyul
Bitta gen, bitta ferment gipotezasi 1940-yillarning boshlarida ilgari surilgan g'oya bo'lib, har bir gen bitta fermentning sintezi yoki faolligini nazorat qiladi. Genetika va biokimyo sohalarini birlashtirgan kontseptsiyani amerikalik genetik Jorj Uells Bidl va Neurospora crassa ustida tadqiqot olib borgan amerikalik biokimyogar Edvard L. Tatum taklif qilgan. Ularning eksperimentlari avval mutatsiyani keltirib chiqaradigan rentgen nurlarining shaklini tasvirlash va keyin uni yovvoyi turdagi shtammning omon qolishi uchun zarur bo'lgan faqat muhim oziq moddalarni o'z ichiga olgan minimal o'sish muhitida etishtirishdan iborat edi. Ular mutant mog'or shtammlari o'sishi uchun ma'lum aminokislotalarni qo'shishni talab qilishini aniqladilar. Ushbu ma'lumotlardan foydalanib, tadqiqotchilar ba'zi genlardagi mutatsiyalarni odatda etishmayotgan aminokislotalarni ishlab chiqaradigan metabolik yo'llardagi o'ziga xos fermentlarning buzilishi bilan bog'lashlari mumkin edi. Endi ma'lumki, barcha genlar fermentni kodlamaydi va ba'zi fermentlar ikki yoki undan ortiq genlar tomonidan kodlangan bir nechta qisqa polipeptidlardan iborat.
Eukaryotik genlarning ekzon-intron tashkilotining kashfiyoti va muqobil qo'shilish imkoniyati shuni ko'rsatdiki, birlamchi transkriptning bir xil nukleotidlar ketma-ketligi bir nechta polipeptid zanjirlarining sintezini ta'minlashi mumkin. turli funktsiyalar yoki ularning o'zgartirilgan analoglari. Misol uchun, xamirturush mitoxondriyalarida nafas olish fermenti sitoxrom b ni kodlaydigan quti (yoki kob) geni mavjud. U ikki shaklda mavjud bo'lishi mumkin: 6400 bp dan iborat "uzun" gen, umumiy uzunligi 1155 bp bo'lgan 6 ta ekzonga ega. va 5 intron. Genning qisqa shakli 3300 bp dan iborat. va 2 intronga ega. Bu aslida birinchi uchta intronga ega bo'lmagan "uzoq" gen. Genning ikkala shakli ham bir xil darajada yaxshi ifodalangan.
Birinchi ikkita ekzonning birlashtirilgan nukleotidlar ketma-ketligiga va ikkinchi intron nukleotidlarining bir qismiga asoslangan "uzun" quti genining birinchi introni olib tashlanganidan so'ng, mustaqil protein - RNK maturaza uchun matritsa hosil bo'ladi (1-rasm). 3.43). RNK maturazasining vazifasi splaysning keyingi bosqichini ta'minlashdan iborat - ikkinchi intronni birlamchi transkriptdan olib tashlash va oxir-oqibat sitoxrom b uchun shablonni shakllantirish.
Yana bir misol - limfotsitlardagi antikor molekulalarining tuzilishini kodlovchi birlamchi transkriptning qo'shilish naqshining o'zgarishi. Antikorlarning membrana shakli C-terminusda aminokislotalarning uzun "dumi" ga ega bo'lib, bu membranada oqsilning mahkamlanishini ta'minlaydi. Antikorlarning ajraladigan shakli bunday quyruqga ega emas, bu esa birlamchi transkripsiyadan bu hududni kodlaydigan nukleotidlarni birlashma paytida olib tashlash bilan izohlanadi.
Viruslar va bakteriyalarda bir gen bir vaqtning o'zida boshqa genning bir qismi bo'lishi mumkin bo'lgan vaziyat tasvirlangan yoki ma'lum bir DNK nukleotid ketma-ketligi ikki xil bir-biriga o'xshash genlarning bir qismi bo'lishi mumkin. Masalan, FX174 fag genomining fizik xaritasi (3.44-rasm) B genining ketma-ketligi A genining ichida joylashganligini va E geni D genining ketma-ketligining bir qismi ekanligini ko'rsatadi. Fagning tashkil etilishining bu xususiyati genom o'zining nisbatan kichik o'lchamlari (u 5386 nukleotiddan iborat) va barcha sintez qilingan oqsillardagi aminokislotalar qoldiqlari soni o'rtasidagi mavjud tafovutni tushuntira oldi, bu ma'lum bir genom sig'imi uchun nazariy jihatdan ruxsat etilganidan oshadi. Bir-biriga o'xshash genlardan (A va B yoki E va D) sintez qilingan mRNKda turli xil peptid zanjirlarini yig'ish imkoniyati ushbu mRNKda ribosomalarni bog'lash joylari mavjudligi bilan ta'minlanadi. Bu boshqa peptidning tarjimasini yangi boshlang'ich nuqtadan boshlash imkonini beradi.
B genining nukleotidlar ketma-ketligi bir vaqtning o'zida A genining bir qismidir va E geni D genining bir qismidir.
l fag genomida ham ramka o'zgarishi bilan va bir xil o'qish ramkasida tarjima qilingan bir-biriga o'xshash genlar topilgan. Bundan tashqari, bitta DNK bo'limining ikkala to'ldiruvchi zanjiridan ikki xil mRNKni transkripsiya qilish mumkin deb taxmin qilinadi. Bu DNK molekulasi bo'ylab turli yo'nalishlarda RNK polimeraza harakatini aniqlaydigan promotor hududlarning mavjudligini talab qiladi.
Bir xil DNK ketma-ketligidan turli xil ma'lumotlarni o'qishga ruxsat etilganligini ko'rsatadigan ta'riflangan holatlar, bir-biriga o'xshash genlar viruslar va, ehtimol, prokaryotlar genomini tashkil qilishning juda keng tarqalgan elementi ekanligini ko'rsatadi. Eukariotlarda genlarning uzilishi bir xil DNK ketma-ketligidan turli xil peptidlarni sintez qilish imkonini beradi.
Bularning barchasini hisobga olgan holda, gen ta'rifiga o'zgartirish kiritish kerak. Shubhasiz, biz endi gen haqida ma'lum bir oqsilni noyob tarzda kodlaydigan DNKning uzluksiz ketma-ketligi haqida gapira olmaymiz. Ko'rinishidan, hozirgi vaqtda "Bir gen - bitta polipeptid" formulasi hali ham eng maqbul deb hisoblanishi kerak, garchi ba'zi mualliflar uni o'zgartirishni taklif qilsalar ham: "Bir polipeptid - bitta gen". Har qanday holatda, gen atamasi o'zining kimyoviy tabiatiga ko'ra polinukleotid bo'lgan va polipeptid zanjiri, tRNK yoki rRNKni sintez qilish imkoniyatini belgilaydigan irsiy materialning funktsional birligi sifatida tushunilishi kerak.
Bitta gen, bitta ferment.
1940-yilda J.Bidl va Edvard Tatum yanada qulayroq tadqiqot mavzusi - mikroskopik qo'ziqorin Neurospora crassada genlarning metabolizmni qanday ta'minlashini o'rganish uchun yangi yondashuvni qo'lladilar.Ular mutatsiyalarni oldilar; u yoki bu metabolik fermentning faolligi yo'q edi. Va bu mutant qo'ziqorin o'z-o'zidan ma'lum bir metabolitni (masalan, leytsin aminokislotasini) sintez qila olmasligiga olib keldi va faqat ozuqaviy muhitga leysin qo'shilganda yashashi mumkin edi. J.Bidl va E.Tatum tomonidan tuzilgan “bir gen, bir ferment” nazariyasi tezda genetiklar orasida keng e’tirofga sazovor bo‘ldi va ularning o‘zlari Nobel mukofotiga sazovor bo‘ldilar.
Usullari. Turli xil metabolik yo'llarni ta'minlaydigan fermentlar ta'sirining buzilishiga olib keladigan "biokimyoviy mutatsiyalar" ni tanlash nafaqat fan, balki amaliyot uchun ham juda samarali bo'ldi. Birinchidan, ular genetika va sanoat mikroorganizmlari seleksiyasining paydo bo'lishiga olib keldi, keyin esa antibiotiklar, vitaminlar, aminokislotalar va boshqalar kabi strategik muhim moddalarni ortiqcha ishlab chiqaradigan mikroorganizmlar shtammlaridan foydalanadigan mikrobiologiya sanoatiga olib keldi.Selektsiya va genetik muhandislik tamoyillari. Superproduser shtammlari "bitta gen bitta fermentni kodlaydi" degan fikrga asoslanadi. Garchi bu g'oya amaliyot uchun juda yaxshi bo'lsa-da, ko'p millionli foyda keltiradi va millionlab odamlarning hayotini (antibiotiklarni) saqlaydi - bu yakuniy emas. Bitta gen faqat bitta ferment emas.
| " |
Genetika- fan hech qanday yosh emas, u bo'yicha tadqiqotlar 1865 yilda Mendeldan to hozirgi kungacha bir necha asrlar davomida olib borildi. Irsiy xususiyat birligi uchun "gen" atamasi birinchi marta 1911 yilda Iogannsen tomonidan taklif qilingan va 1940-yillarda Tatum va Bidl tomonidan taklif qilingan "bir gen, bitta ferment" kontseptsiyasi tomonidan aniqlangan.
Bu pozitsiya Drosophila chivinlari ustida o'tkazilgan tajribalarda aniqlangan, ammo odamlar uchun ham xuddi shunday; Oxir oqibat, barcha mavjudotlarning hayoti ularning DNKsi bilan belgilanadi. Inson DNK molekulasi boshqa barcha organizmlarnikidan kattaroq bo'lib, tuzilishi murakkabroq, lekin uning funktsiyalarining mohiyati barcha tirik mavjudotlarda bir xildir.
Kontseptsiya " bitta gen - bitta ferment Tatum va Beadl g'oyalari asosida paydo bo'lgan ", quyidagicha ifodalanishi mumkin:
1. Hamma narsa biologik jarayonlar genetik nazorat ostida.
2. Barcha biokimyoviy jarayonlar bosqichma-bosqich reaktsiyalar shaklida sodir bo'ladi.
3. Har bir biokimyoviy reaksiya oxir-oqibatda turli individual genlar nazorati ostida bo‘ladi.
4. Muayyan gendagi mutatsiya hujayraning ma'lum bir kimyoviy reaktsiyani amalga oshirish qobiliyatining o'zgarishiga olib keladi.
O'shandan beri "bitta gen - bitta ferment" tushunchasi biroz kengaydi va endi " bitta gen - bitta protein" Bundan tashqari, yaqinda o'tkazilgan tadqiqotlar shuni ko'rsatadiki, ba'zi genlar boshqalar bilan birgalikda noyob oqsillarni ishlab chiqarish uchun harakat qiladi, ya'ni ba'zi genlar bir nechta oqsillarni kodlashi mumkin.
Inson genomi 3 milliardga yaqin nukleotid juftlarini o'z ichiga oladi; unda 50 000 dan 100 000 gacha bo'lganligi taxmin qilinadi. Genomni deshifrlashdan so‘ng bor-yo‘g‘i 30 mingga yaqin gen borligi ma’lum bo‘ldi.Bu genlarning o‘zaro ta’siri kutilganidan ancha murakkabroq. Genlar DNK zanjirlarida kodlangan bo'lib, ular ma'lum yadro oqsillari bilan xromosomalarni hosil qiladi.
Genlar- shunchaki DNK segmentlari emas: ular kodlash ketma-ketliklari - ekzonlar, kodlanmaydigan ketma-ketliklar - nitronlar bilan kesishgan holda hosil bo'ladi. Ekzonlar DNKning ifodalangan qismi sifatida organizmning eng muhim molekulasining faqat kichik qismini tashkil qiladi; uning katta qismi ifodalanmaydi, nitronlar tomonidan hosil bo'ladi va ko'pincha "jim" DNK deb ataladi.
Taxminan hajmi va tuzilishi inson genomi quyidagi rasmda keltirilgan. Inson xromosomasining funksional uzunligi sentimorganidlarda ifodalanadi. Centimorganide (cM) - bu meyoz paytida kesishish ehtimoli 1% bo'lgan masofa. Gen aloqasi tahlili shuni ko'rsatdiki, inson genomining uzunligi taxminan 3000 sm.
O'rtacha xromosoma 130 million nukleotid asos juftlarida kodlangan taxminan 1500 genni o'z ichiga oladi. Quyidagi rasmda genomning jismoniy va funktsional o'lchamlari sxematik tarzda ko'rsatilgan: birinchisi nukleotid juftlarida, ikkinchisi esa cMda hisoblanadi. Inson genomining ko'p qismi "jim" DNK bilan ifodalanadi va ifoda etilmaydi.
Yoniq DNK matritsasi Transkripsiya jarayoni natijasida RNK, keyin esa oqsil sintezlanadi. Binobarin, DNK ketma-ketligi hujayraning funktsional oqsillari ketma-ketligini to'liq aniqlaydi. Barcha oqsillar quyidagicha sintezlanadi:
DNK => RNK => oqsil

Odamlar va boshqa sutemizuvchilarning genetik apparati boshqa tirik organizmlarga qaraganda ancha murakkab, chunki sutemizuvchilardagi ba'zi genlarning bo'limlari boshqalarining qismlari bilan birlashtirilishi mumkin. genlar, natijada butunlay yangi oqsil sintezi yoki alohida hujayra funktsiyasini nazorat qilish.
Binobarin, odamlarda ifodalangan genlar hajmini amalda ko'paytirmasdan ko'paytirish mumkin. DNK yoki genlarning mutlaq soni.
Umuman olganda, barcha genetik materialning taxminan 70% ifodalanmaydi.
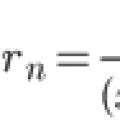 Teylor qatorini kengaytirish Koshi muammosining oddiy uchun taxminiy yechimi
Teylor qatorini kengaytirish Koshi muammosining oddiy uchun taxminiy yechimi Maktabda nima o'rgatilmaydi, maktabda nima o'rgatilmaydi, deb so'rang
Maktabda nima o'rgatilmaydi, maktabda nima o'rgatilmaydi, deb so'rang Pulni o'ylash formulasi (A
Pulni o'ylash formulasi (A