Struktur og biologisk rolle av nukleotider, nukleinsyrer. DNA-replikasjon og transkripsjon
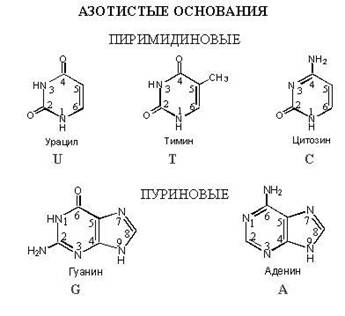
Nukleotider er fosforestere av nukleosider.
Deres kjemiske sammensetning: nitrogenholdig base (A.O.) + pentose + fosforsyre
Fosforestere dannes med deltagelse av hydroksylgrupper av pentoser. Posisjonene til fosforestergrupper er vanligvis utpekt ved å bruke notasjonen ("), for eksempel: 5" , 3 "
Innledende kort informasjon: Nukleotider spiller en ekstremt viktig rolle i cellens liv.
Klassifisering av nukleotider
Nukleotider som består av ett molekyl A.O, pentose, fosforsyre, er kalt mononukleotider. Mononukleotider kan inneholde ett molekyl fosforsyre, to eller tre molekyler fosforsyre koblet til hverandre.
Kombinasjon av to mononukleotider vanligvis kalt dinukleotid. I Dinukleotidet inneholder vanligvis forskjellige nitrogenholdige baser eller en annen syklisk forbindelse, for eksempel et vitamin.
Sykliske mononukleotider spiller en spesiell rolle i biokjemiske prosesser.
Nomenklatur av mononukleotider.
Til tittelen nukleosid lagt til basert på mengden fosfatrester, '' monofosfatʼʼ, ʼʼdifosfatʼʼ, "trifosfat"ʼʼ, som indikerer deres posisjon i pentosesyklusen - digital betegnelse av stedet med et symbol ("),
Plasseringen av fosfatgruppen i posisjon (5") er den vanligste og typiske, derfor kan den utelates (AMP, GTP, UTP, d AMF, etc.)
De resterende posisjonene må angis (3" - AMP, 2" - AMP, 3" - d AMF)
5"-adenosinmonofosfat
(5" - AMF eller AMF)
Navn på de vanligste nukleotidene
| nukleosid | nukleosidmonofosfat | nukleosid-difosfat | nukleosidtrifosfat |
| adenosin | 5"-adenosinmonofosfat (5"-AMP eller AMP) 5"-adenylsyre | 5"-adenosindifosfat (5"-ADP eller ADP) | 5"-adenosintrifosfat (5"-ATP eller ATP) |
| adenosin | 3"-adenosinmonofosfat (3"-AMP) 3"-adenylsyre | ikke funnet in vivo | ikke funnet in vivo |
| guanosin | 5"-guanosinmonofosfat (5"-GMP eller GMP) | 5"-guanosin difosfat (5"-BNP eller HDF) | 5"-guanosintrifosfat (5"-GTP eller GTP) |
| guanosin | 3"-guanosinmonofosfat (3"-GMP) 3"-guanylsyre | ikke funnet in vivo | ikke funnet in vivo |
| deoksy adenosin | 5"-deoksyadenosinmonofosfat (5"- d AMF eller d AMF) | 5"-deoksyadenosindifosfat (5"- d ADFili d ADF) | 5"-deoksyadenosintrifosfat (5"- d ATFili d ATP) |
| uridin | 5"-uridinmonofosfat (5"-UMP eller UMP) | 5"-uridin difosfat (5"-UDP eller UDF) | 5"-uridintrifosfat (5"-UTP eller UTP) |
| cytidin | 5"-cytidinmonofosfat (5"- CMP eller CMP) | 5"-cytidin difosfat (5"-CDP eller CDP) | 5"-cytidintrifosfat (5"-CTP eller CTP) |
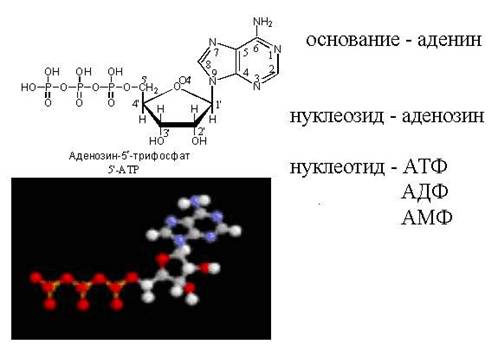
Nukleotider dannet med deltakelse av ribose kan inneholde fosforsyrerester i tre posisjoner (5", 3", 2"), og med deltakelse av deoksyribose - bare i to posisjoner (5", 3"), i posisjon 2" der er ingen hydroksygruppe.Denne omstendigheten er svært viktig for strukturen til DNA.
Fraværet av en hydroksygruppe i den andre posisjonen har to viktige konsekvenser:
Polarisasjonen av glykosidbindingen i DNA avtar og den blir mer motstandsdyktig mot hydrolyse.
2-O-deoksyribose kan ikke gjennomgå verken epimerisering eller omdannelse til ketose.
I cellen blir nukleosidmonofosfat suksessivt omdannet til difosfat og deretter til trifosfat.
For eksempel: AMP ---> ADP ---> ATP
Biologisk rolle til nukleotider
Alle nukleosid-difosfater Og nukleosidtrifosfater tilhører høyenergiske (makroerge) forbindelser.
Nukleosidtrifosfater delta i syntese nukleinsyrer, sikre aktivering av bioorganiske forbindelser og biokjemiske prosesser som skjer med energiforbruk. Adenosintrifosfat (ATP) er den vanligste høyenergiforbindelsen i menneskekroppen. ATP-innholdet i skjelettmuskulaturen til pattedyr er opptil 4 g/kg, generelt innhold ca 125 ᴦ. Hos mennesker når hastigheten på ATP-metabolisme 50 kg/dag. Hydrolysen av ATP produserer adenosindifosfat(ADF)
Makroergiske forbindelser
ATP inneholder forskjellige typer kjemiske bindinger:
N-β-glykosid
Ester
To anhydrider (biologisk høyenergi)
I forhold in vivo hydrolyse av høyenergi-ATP-bindingen er ledsaget av frigjøring av energi (ca. 35 kJ/mol), som gir andre energiavhengige biokjemiske prosesser.
ATP + H2O - enzym ATP hydrolase -> ADP + H3 PO4
I vandige løsninger ADP og ATP ustabil . Ved 0 0 er SATP stabil i vann i bare noen få timer, og ved koking i 10 minutter.
Under påvirkning av alkali hydrolyseres de to terminale fosfatene (anhydridbindinger) lett, men den siste (esterbindingen) er vanskelig. Under syrehydrolyse blir N-glykosidbindingen lett ødelagt.
For første gang ble ATP frigjort fra muskler i 1929 ᴦ. K. Loman. Kjemisk syntese ble utført i 1948 ᴦ. A. Todd.
Sykliske nukleotider er mellomledd i overføringen av hormonsignaler, og endrer aktiviteten til enzymer i cellen.
Οʜᴎ dannes fra nukleosidtrifosfater.
ATP - cyclase enzym -> cAMP + H4 P2 O7
Etter at handlingen er fullført, oppstår hydrolyse av det sykliske nukleotidet. . To forbindelser, 5"-AMP og 3"-AMP, kan dannes, men under biologiske forhold dannes det bare 5"-AMP,
Syklisk adenosinmonofosfat (cAMP)
11.5.Struktur av nukleinsyrer
Den primære strukturen til RNA og DNA er den sekvensielle forbindelsen av nukleotider i en polynukleotidkjede. Skjelettet til en polynukleotidkjede består av karbohydrat- og fosfatrester; heterosykliske nitrogenholdige baser er koblet til karbohydrater gjennom N-β glykosidbindinger. Fra et biologisk synspunkt er de viktigste tripletter – blokker av nukleotider som består av tre nitrogenholdige baser, som hver koder for en aminosyre eller har en spesifikk signalfunksjon.
Strukturen til NC kan representeres skjematisk:
5" 3" 5" 3" 5" 3"
fosfat -- pentose -- fosfat -- pentose -- fosfat -- pentose -OH
I den primære strukturen til DNA Start kjeder bestemmes av en pentose som inneholder et fosfat i posisjon 5". Pentoser i en polynukleotidkjede er forbundet gjennom fosfatbindinger 3 "→ 5". På slutt kjeder i posisjon 3"-pentose OH-gruppe forblir frie.
Høyere ordens DNA-struktur - dobbel helix
Vitenskapelig beskrivelse sekundær struktur DNA refererer til største funn menneskeheten i det tjuende århundre. Biokjemiker D. Watson og fysiker F. Crick i 1953 foreslo de en modell av strukturen til DNA og mekanismen for replikasjonsprosessen. I 1962. de ble tildelt Nobelprisen.
I sin populære form er historien beskrevet i James Watsons bok "The Double Helix", M.: Mir, 1973. Boken beskriver veldig interessant samarbeidets historie, med humor og lett ironi av forfatteren for slikt betydelig hendelse, hvor de lykkelige "skyldige" var to unge forskere. Siden oppdagelsen av DNA-strukturen har menneskeheten fått et verktøy for utvikling av en ny retning - bioteknologi, proteinsyntese gjennom genrekombinasjon (hormoner i medisinsk industri produseres av insulin, erytropoietin og mange andre).
Forskning bidro til oppdagelsen av strukturen til DNA E. Chargaffa om den kjemiske sammensetningen av DNA. Han fant ut:
Antall pyrimidinbaser er lik antallet purinbaser
Mengden tymin er lik mengden adenin, og mengden cytosin er lik
A = T G = C
A + G = T + C
A + C = T + G
Dette forholdet kalles Chargaff regler .
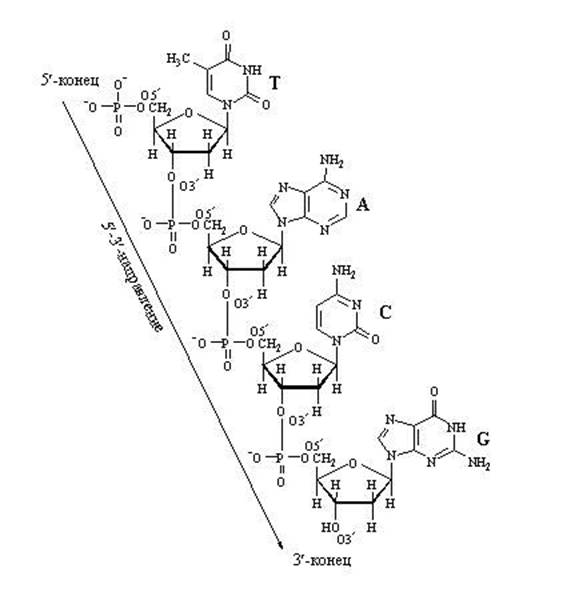
DNA-molekylet består av to vridde helixer. Skjelettet til hver helix er en kjede av vekslende deoksyribose- og fosforsyrerester. Spiralene er orientert på en slik måte at de danner to ulike spiralspor som går parallelt med hovedaksen. Disse sporene er fylt med proteiner histoner. Nitrogenbasene er plassert inne i helixen, nesten vinkelrett på hovedaksen og danner komplementære par mellom kjedene A...T og G...C.
Den totale lengden på DNA-molekylene i hver celle når 3 cm. Cellediameteren er i gjennomsnitt 10–5 m, DNA-diameteren er bare 2‣‣‣10–9 m.
Grunnleggende parametere for den doble helixen:
* diameter 1,8 – 2 nm,
* en tur inneholder 10 nukleotider
* svinghøyde ~ 3,4 nm
* avstanden mellom to nukleotider er 0,34 nm.
Basene er plassert vinkelrett på kjedens akse.
* retningene til polynukleotidkjeder er antiparallelle
* forbindelse mellom furanose sykluser av deoksyribose via
fosforsyre utføres fra posisjon 3` til posisjon 5` in
hver av kretsene.
* Begynnelsen av kjeden – hydroksylgruppen til pentose er fosforylert i posisjon
5', er enden av kjeden den frie hydroksylgruppen til pentose i posisjon 3'.
* I DNA og RNA er nukleosidfragmenter i antikonformasjon; pyrimidinringen til purin er lokalisert til høyre for glykosidbindingen. Bare denne posisjonen tillater dannelse av et komplementært par (se nukleotidformler)
* Tre typer interaksjoner forekommer mellom nitrogenholdige baser:
1. "Tverrgående", komplementære par av to kjeder er involvert. En "syklisk" overføring av elektroner skjer mellom to nitrogenholdige baser (T - A, U - C), et ekstra p-elektronsystem dannes, som gir ytterligere interaksjon og beskytter nitrogenholdige baser mot uønsket kjemisk påvirkning. Mellom to hydrogenbindinger etableres mellom adenin og tymin, og tre hydrogenbindinger mellom guanin og cytosin.
2. "Vertikal" (stabling), på grunn av stabling, er de nitrogenholdige basene i en kjede involvert. "Stable interaction" har til og med mer betydning for å stabilisere strukturen enn samspillet i komplementære par
3. Interaksjon med vann spiller en betydelig rolle i å opprettholde den romlige strukturen til dobbelthelixen, som tar på seg den mest kompakte strukturen for å redusere overflaten av kontakt med vann og leder hydrofobe heterosykliske baser inn i helixen.
Struktur og sammensetning av nukleoproteinkomplekser
Flere typer interaksjoner deltar i bindingen av nukleinsyre til protein:
Elektrostatisk
Hydrogenbindinger
Hydrofobisk
Basert på resultatene av røntgenstrukturanalyse ble ekte tredimensjonale modeller av DNA, ribosomer, informasjonsosomer og virale nukleinsyrer konstruert ved bruk av datamodellering.
Histon DNA-proteiner har distinkte grunnleggende egenskaper og kjennetegnes ved en høy grad av evolusjonær bevaring. Basert på forholdet mellom de to grunnleggende aminosyrene lysin/arginin er de delt inn i 5 klasser: H1, H2A, H 2B, H3, H4
Nukleotider - konsept og typer. Klassifisering og funksjoner i kategorien "Nukleotider" 2017, 2018.
- dette er komplekse monomerer som heteropolymermolekyler er satt sammen fra. DNA og RNA. Frie nukleotider er involvert i signal- og energiprosesser i livet. DNA-nukleotider og RNA-nukleotider har en felles strukturplan, men er forskjellige i strukturen til pentosesukkeret. DNA-nukleotider bruker sukkeret deoksyribose, mens RNA-nukleotider bruker ribose.
Nukleotidstruktur
Hvert nukleotid kan deles inn i 3 deler:
1. Et karbohydrat er et femleddet pentosesukker (ribose eller deoksyribose).
2. Fosforrest (fosfat) er en rest av fosforsyre.
3. En nitrogenholdig base er en forbindelse som inneholder mange nitrogenatomer. Nukleinsyrer bruker kun 5 typer nitrogenholdige baser: Adenin, Thymin, Guanin, Cytosin, Uracil. Det er 4 typer i DNA: Adenin, Thymin, Guanin, Cytosin. Det er også 4 typer i RNA: Adenin, Uracil, Guanin, Cytosin Det er lett å legge merke til at i RNA er thymin erstattet med Uracil sammenlignet med DNA.
Generell strukturformel pentose (ribose eller deoksyribose), hvis molekyler danner "skjelettet" av nukleinsyrer:
Hvis X er erstattet med H (X = H), oppnås deoksyribonukleosider; hvis X er erstattet med OH (X = OH), oppnås ribonukleosider. Hvis du erstatter en nitrogenholdig base (purin eller pyrimidin) i stedet for R, får du et spesifikt nukleotid.
Det er viktig å ta hensyn til posisjonene til karbonatomer i pentose, som er betegnet som 3" og 5". Nummereringen av karbonatomer starter fra oksygenatomet øverst og går med klokken. Det siste er et karbonatom (5"), som befinner seg utenfor pentoseringen og danner, kan man si, en "hale" av pentosen. Så når en kjede av nukleotider vokser, kan enzymet bare feste et nytt nukleotid til karbon 3" og ikke til noen andre . Derfor kan 5"-enden av nukleotidkjeden aldri fortsettes; bare 3"-enden kan forlenges.


Sammenlign nukleotidet for RNA med nukleotidet for DNA.
Prøv å finne ut hvilket nukleotid det er i denne representasjonen:

ATP - fritt nukleotid
cAMP er et "løkke" ATP-molekyl

Nukleotidstrukturdiagram
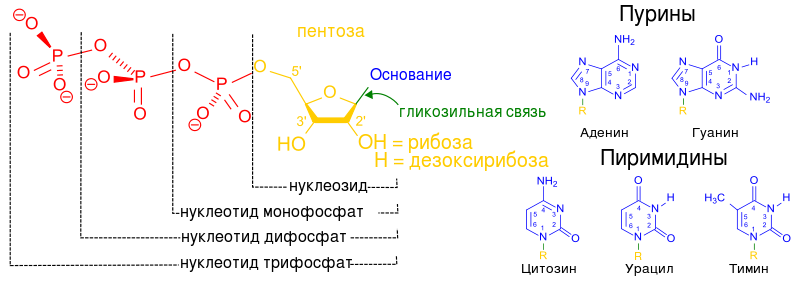
Vær oppmerksom på at det aktiverte nukleotidet, som er i stand til å forlenge DNA- eller RNA-kjeden, har en "trifosfathale". Det er med denne "energimettede" halen at den kan slutte seg til en eksisterende kjede av voksende nukleinsyre. Fosfathalen sitter på 5. karbon, så denne karbonposisjonen er allerede okkupert av fosfater og er der for festing. Hva skal jeg feste den til? Bare til karbonet i posisjon 3". Når det først er festet, vil et gitt nukleotid i seg selv bli et mål for bindingen til det neste nukleotidet. Den "mottakende siden" gir karbonet i posisjon 3", og den "ankommende siden" klamrer seg til det med fosfathalen plassert i posisjon 5". Generelt vokser kjeden fra 3"-siden.
Forlengelse av DNA-nukleotidkjeden
Kjedeforlengelsen på grunn av "langsgående" bindinger mellom nukleotider kan bare gå i én retning: fra 5" ⇒ til 3", fordi et nytt nukleotid kan bare legges til 3"-enden av kjeden, men ikke til 5"-enden.
Par av nukleotider forbundet med komplementære tverrbindinger av deres nitrogenholdige baser
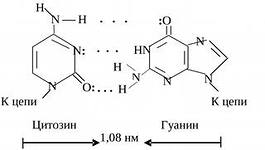

DNA dobbel helix region

Finn tegn på at to DNA-tråder er antiparallelle.
Finn nukleotidpar med doble og trippel komplementære bindinger.
Nukleosider er på sin side N-glykosider som inneholder en heterosyklisk del koblet via et nitrogenatom til C-1-atomet i en sukkerrest.
I naturen er de vanligste nukleotidene β-N-glykosider av puriner eller pyrimidiner og pentoser - D-ribose eller D-2-deoksyribose. Avhengig av strukturen til pentose skilles det mellom ribonukleotider og deoksyribonukleotider, som er monomerer av molekyler av komplekse biologiske polymerer (polynukleotider)- henholdsvis RNA eller DNA.
Fosfatresten i nukleotidene danner vanligvis en esterbinding med 2'-, 3'- eller 5'-hydroksylgruppene til ribonukleosidene; i tilfellet med 2'-deoksynukleosidene er 3'- eller 5'-hydroksylgruppene forestret.
De fleste nukleotidene er monoestere av ortofosforsyre, men det er også kjent diestere av nukleotider der to hydroksylrester er forestret - for eksempel de sykliske nukleotidene cykloadenin og cykloguaninmonofosfater (cAMP og cGMP). Sammen med nukleotider - estere av ortofosforsyre (monofosfater), mono- og diestere av pyrofosforsyre (difosfater, for eksempel adenosindifosfat) og monoestere av tripolyfosforsyre (trifosfater, for eksempel adenosintrifosfat) er også vanlige i naturen.
Nomenklatur
| Kode | Midler | Komplementært par |
|---|---|---|
| EN | T i DNA; U i RNA |
|
| C | G | |
| G | C | |
| eller |
T i DNA; U i RNA |
EN |
| M | EN eller C |
K |
| R | EN eller G |
Y |
| W | EN eller T |
W |
| S | C eller G |
S |
| Y | C eller T |
R |
| K | G eller T |
M |
| V | EN eller C eller G |
B |
| H | EN eller C eller T |
D |
| D | EN eller G eller T |
H |
| B | C eller G eller T |
V |
| X eller N |
EN eller C eller G eller T(U) |
noen |
Forbindelser som består av to nukleotidmolekyler kalles dinukleotider, av tre - trinukleotider, fra et lite antall - oligonukleotider, og fra mange - polynukleotider eller nukleinsyrer.
Navnene på nukleotider er forkortelser i form av standard tre- eller firebokstavskoder.
Hvis forkortelsen begynner med en liten bokstav "d" (eng. d), som betyr deoksyribonukleotid; fraværet av bokstaven "d" betyr ribonukleotid. Hvis forkortelsen begynner med en liten bokstav "ts" (eng. c), som betyr at vi snakker om den sykliske formen til nukleotidet (for eksempel cAMP).
Den første store bokstaven i forkortelsen indikerer en spesifikk nitrogenholdig base eller gruppe av mulige nukleinbaser, den andre bokstaven indikerer antall fosforsyrerester i strukturen (M - mono-, D - di-, T - tri-), og den tredje store bokstaven er alltid bokstaven F ("-fosfat"; engelsk. P).
Latinske og russiske koder for nukleisk baser:
- T - T: Tymin (5-metyluracil), funnet i bakteriofager i DNA, tar plassen til uracil i RNA;
- U - U: Uracil, som finnes i RNA, tar plassen til tymin i DNA.
De generelt aksepterte bokstavkodene for å betegne nukleotidbaser tilsvarer nomenklaturen vedtatt av International Union of Pure and Applied Chemistry (forkortet engelsk). IUPAC, IUPAC) og International Union of Biochemistry and Molecular Biology (Engelsk)russisk, forkortet - engelsk. IUBMB). Hvis det ved sekvensering av en DNA- eller RNA-sekvens er tvil om nøyaktigheten av å bestemme et bestemt nukleotid, i tillegg til de fem viktigste (A, C, T, G, U), brukes andre bokstaver i det latinske alfabetet avhengig av på hva de mest sannsynlige nukleotidene kan være i en gitt sekvensposisjon. De samme tilleggsbokstavene brukes for å angi degenererte posisjoner (som ikke sammenfaller i forskjellige homologe sekvenser), for eksempel når du skriver sekvensen av primere for PCR.
Lengden på sekvenserte DNA-seksjoner (gen, sted, kromosom) eller hele genomet er indikert i nukleotidpar (bp) eller basepar (eng. basepar, forkortet bp), som med dette betyr den elementære enheten til et dobbelttrådet nukleinsyremolekyl, sammensatt av to parede komplementære baser.
Biologisk rolle
1. Universell energikilde (ATP og dets analoger).
2. De er aktivatorer og bærere av monomerer i cellen (UDP-glukose)
6. De er monomerer i sammensetningen av nukleinsyrer, forbundet med 3′-5′-fosfodiesterbindinger.
Historie
I premolekylær genetikk ble et spesielt begrep brukt for å betegne det minste elementet i DNA-strukturen som kan bli utsatt for spontan eller indusert mutasjon rekon. Det har nå vist seg at det minste grunnstoffet er ett nukleotid (eller en nitrogenholdig base i et nukleotid), så dette begrepet brukes ikke lenger. For å definere konseptet mutasjonsenhet begrepet ble brukt Mouton. Det er nå vist at en mutasjon kan manifestere seg fenotypisk selv når et enkelt nukleotid (eller nitrogenholdig base i et nukleotid) erstattes, og dermed tilsvarer begrepet muton et enkelt nukleotid.
Skriv en anmeldelse om artikkelen "Nukleotider"
Notater
se også
Linker
- Nukleotider- artikkel fra Great Soviet Encyclopedia.
- Bokstavkoder for nukleotidbaser i henhold til IUPAC- og IUBMB-nomenklaturen (Engelsk)russisk
- i nettmagasinet
|
||||||||||||||||||||||||||||||||||
Utdrag som karakteriserer nukleotider
Kosaken ble oppringt og avhørt; Kosakkkommandantene ønsket å benytte anledningen til å gjenerobre hestene, men en av befalene, kjent med de høyeste gradene i hæren, rapporterte dette faktum til stabsgeneralen. Situasjonen ved hærens hovedkvarter har den siste tiden vært ekstremt spent. Ermolov, noen dager før, etter å ha kommet til Bennigsen, tryglet ham om å bruke sin innflytelse på øverstkommanderende for å få til en offensiv."Hvis jeg ikke kjente deg, ville jeg tro at du ikke vil ha det du ber om." "Så snart jeg råder én ting, vil Hans fredelige høyhet sannsynligvis gjøre det motsatte," svarte Bennigsen.
Nyheten om kosakkene, bekreftet av de sendte patruljene, beviste hendelsens endelige modenhet. Den strakte strengen hoppet, og klokken hveste og klokkespillet begynte å spille. Til tross for all hans imaginære kraft, hans intelligens, erfaring, kunnskap om mennesker, Kutuzov, tatt i betraktning notatet fra Bennigsen, som personlig sendte rapporter til suverenen, det samme ønsket uttrykt av alle generalene, ønsket til suverenen overtatt av ham og sammenføringen av kosakkene, kunne ikke lenger begrense uunngåelig bevegelse og ga ordre om det han anså som ubrukelig og skadelig - han velsignet det fullførte faktum.
Notatet Bennigsen sendte inn om behovet for en offensiv, og informasjonen fra kosakkene om franskmennenes avdekkede venstre flanke var bare de siste tegnene på behovet for å bestille en offensiv, og offensiven var berammet til 5. oktober.
Om morgenen den 4. oktober undertegnet Kutuzov disposisjonen. Tol leste den for Yermolov og inviterte ham til å ta seg av ytterligere bestillinger.
"Ok, ok, jeg har ikke tid nå," sa Ermolov og forlot hytta. Den disposisjonen Tol utarbeidet var meget god. Akkurat som i Austerlitz-disposisjonen ble det skrevet, men ikke på tysk:
"Die erste Colonne marschiert [den første kolonnen går (tysk)] denne veien og den, die zweite Colonne marschiert [den andre kolonnen går (tysk)] denne veien og den veien," osv. Og alle disse spaltene på papiret kom de til deres plass til fastsatt tid og ødela fienden. Alt var, som i alle disposisjoner, perfekt gjennomtenkt, og som i alle disposisjoner kom ikke en eneste kolonne til sin tid og på sin plass.
Da disposisjonen var klar i det nødvendige antall eksemplarer, ble en offiser tilkalt og sendt til Ermolov for å gi ham papirene for henrettelse. En ung kavalerioffiser, Kutuzovs ordfører, fornøyd med viktigheten av oppdraget som ble gitt ham, dro til Ermolovs leilighet.
«Vi har dratt,» svarte Yermolovs ordinære. Kavalerioffiseren dro til generalen, som ofte besøkte Ermolov.
– Nei, og det er ingen generell.
Kavalerioffiseren, sittende på hesteryggen, red til en annen.
– Nei, de dro.
«Hvordan kunne jeg ikke være ansvarlig for forsinkelsen! Så synd! - tenkte betjenten. Han turnerte hele leiren. Noen sa at de så Ermolov gå et sted sammen med andre generaler, noen sa at han sannsynligvis var hjemme igjen. Betjenten, uten å spise lunsj, lette frem til klokken seks om kvelden. Yermolov var ingensteds og ingen visste hvor han var. Offiseren tok raskt en matbit med en kamerat og gikk tilbake til fortroppen for å se Miloradovich. Miloradovich var heller ikke hjemme, men da ble han fortalt at Miloradovich var på general Kikins ball, og at Yermolov måtte være der også.
- Hvor er det?
"Der borte, i Echkino," sa kosakkoffiseren og pekte på en fjerntliggende grunneiers hus.
– Hvordan er det der, bak kjedet?
– De sendte to av regimentene våre inn i en kjede, det foregår en slik fest der nå, det er en katastrofe! To musikk, tre kor med låtskrivere.
Offiseren gikk bak lenken til Echkin. På lang avstand, nærmet seg huset, hørte han de vennlige, muntre lydene av en soldats dansende sang.
“I engene, ah... in the meadows!...” - hørte han ham plystre og klirre, av og til overdøvet av rop fra stemmer. Offiseren følte seg glad i sjelen av disse lydene, men samtidig var han redd for at han var skyld i at han ikke overførte den viktige ordren som var betrodd ham så lenge. Klokken var allerede ni. Han steg av hesten og gikk inn på verandaen og inngangspartiet til en stor, intakt herregård, som ligger mellom russerne og franskmennene. I pantryet og i gangen tudlet det rundt med vin og matretter. Det var sangbøker under vinduene. Offiseren ble ført gjennom døren, og han så plutselig alle de viktigste generalene i hæren sammen, inkludert den store, merkbare skikkelsen til Ermolov. Alle generalene var i uknappede frakker, med røde, livlige ansikter og lo høyt, og sto i en halvsirkel. Midt i salen var en kjekk kort general med rødt ansikt i ferd med å lage en thrasher smart og behendig.
– Ha, ha, ha! Å ja Nikolai Ivanovich! ha, ha, ha!..
Offiseren følte at ved å gå inn i dette øyeblikk med en viktig ordre, var han dobbelt skyldig, og han ville vente; men en av generalene så ham, og etter å ha lært hva han var for noe, fortalte han Ermolov. Ermolov, med et rynket ansikt, gikk ut til betjenten og etter å ha lyttet tok han avisen fra ham uten å fortelle ham noe.
– Tror du han dro ved et uhell? - sa en stabskamerat til en kavalerioffiser om Ermolov den kvelden. – Dette er ting, alt er med vilje. Gi Konovnitsyn en tur. Se, for et rot det blir i morgen!
Dagen etter, tidlig om morgenen, sto den avfeldige Kutuzov opp, ba til Gud, kledde på seg, og med den ubehagelige bevisstheten at han måtte lede en kamp som han ikke godkjente, satte seg i en vogn og kjørte ut av Letashevka , fem mil bak Tarutin, til stedet hvor de fremrykkende søylene skulle settes sammen. Kutuzov red, sovnet og våknet og lyttet for å se om det var noen skudd til høyre, om ting begynte? Men alt var fortsatt stille. Daggryet til en fuktig og overskyet høstdag var så vidt begynt. Da han nærmet seg Tarutin, la Kutuzov merke til kavalerister som førte hestene sine til vann over veien som vognen kjørte langs. Kutuzov tok en nærmere titt på dem, stoppet vognen og spurte hvilket regiment? Kavaleristene var fra kolonnen som skulle ha vært langt fremme i bakhold. «Det kan være en feil,» tenkte den gamle øverstkommanderende. Men etter å ha kjørt enda lenger, så Kutuzov infanteriregimenter, våpen i boksene, soldater med grøt og ved, i underbukser. En offiser ble tilkalt. Offiseren rapporterte at det ikke var noen ordre om å flytte.
«Hvordan kunne du ikke...» begynte Kutuzov, men ble umiddelbart stille og beordret senioroffiseren å bli kalt til ham. Etter å ha gått ut av vognen, med hodet ned og pustet tungt, stille ventende, gikk han frem og tilbake. Da den forespurte generalstabsoffiseren Eichen dukket opp, ble Kutuzov lilla, ikke fordi denne offiseren gjorde seg skyldig i en feil, men fordi han var et verdig emne for å uttrykke sinne. Og rystende, gisper etter pusten, gikk den gamle mannen inn i den raseriet han var i stand til å gå inn i da han rullet på bakken i sinne, angrep han Eichen, truet med hendene, ropte og sverget i vulgært. ord. En annen person som dukket opp, kaptein Brozin, som var uskyldig i noe, led samme skjebne.
Alt liv på planeten består av mange celler som opprettholder orden i deres organisasjon på grunn av den genetiske informasjonen som finnes i kjernen. Det lagres, implementeres og overføres av komplekse høymolekylære forbindelser - nukleinsyrer, bestående av monomere enheter - nukleotider. Rollen til nukleinsyrer kan ikke overvurderes. Stabiliteten til strukturen deres bestemmer kroppens normale funksjon, og eventuelle avvik i strukturen fører uunngåelig til endringer i cellulær organisasjon, aktiviteten til fysiologiske prosesser og levedyktigheten til celler generelt.
Konseptet med et nukleotid og dets egenskaper
Hvert RNA er satt sammen av mindre monomere forbindelser - nukleotider. Et nukleotid er med andre ord et byggemateriale for nukleinsyrer, koenzymer og mange andre biologiske forbindelser som er essensielle for en celle i løpet av dens levetid.
Hovedegenskapene til disse essensielle stoffene inkluderer:
Lagre informasjon om og arvede egenskaper;
. utøve kontroll over vekst og reproduksjon;
. deltakelse i metabolisme og mange andre fysiologiske prosesser som skjer i cellen.
Når vi snakker om nukleotider, kan man ikke unngå å dvele ved et så viktig spørsmål som deres struktur og sammensetning.

Hvert nukleotid består av:
Sukkerrester;
. nitrogenholdig base;
. fosfatgruppe eller fosforsyrerest.
Vi kan si at et nukleotid er en kompleks organisk forbindelse. Avhengig av artssammensetningen av nitrogenholdige baser og typen pentose i nukleotidstrukturen, er nukleinsyrer delt inn i:
deoksyribonukleinsyre, eller DNA;
. ribonukleinsyre eller RNA.
Nukleinsyresammensetning
I nukleinsyrer er sukker representert med pentose. Det er et fem-karbon sukker, kalt deoksyribose i DNA og ribose i RNA. Hvert pentosemolekyl har fem karbonatomer, fire av dem danner sammen med oksygenatomet en femleddet ring, og det femte er en del av HO-CH2-gruppen.
Plasseringen til hvert karbonatom i pentosemolekylet er indikert med et arabisk tall med et primtall (1C´, 2C´, 3C´, 4C´, 5C´). Siden alle prosesser for å lese fra et nukleinsyremolekyl har en streng retning, tjener nummereringen av karbonatomer og deres plassering i ringen som en slags indikator på riktig retning.
Ved hydroksylgruppen er en fosforsyrerest festet til det tredje og femte karbonatomet (3C´ og 5C´). Den bestemmer den kjemiske tilknytningen av DNA og RNA til gruppen av syrer.
En nitrogenholdig base er festet til det første karbonatomet (1C´) i et sukkermolekyl.
Artssammensetning av nitrogenholdige baser
DNA-nukleotider basert på nitrogenbasen er representert av fire typer:
adenin (A);
. guanin (G);
. cytosin (C);
. tymin (T).
De to første tilhører klassen puriner, de to siste tilhører klassen pyrimidiner. Når det gjelder molekylvekt, er puriner alltid tyngre enn pyrimidiner.

RNA-nukleotider basert på nitrogenbasen er representert av:
adenin (A);
. guanin (G);
. cytosin (C);
. uracil (U).
Uracil, som tymin, er en pyrimidinbase.
I den vitenskapelige litteraturen kan du ofte finne en annen betegnelse for nitrogenholdige baser - med latinske bokstaver (A, T, C, G, U).
La oss dvele mer detaljert på den kjemiske strukturen til puriner og pyrimidiner.

Pyrimidiner, nemlig cytosin, tymin og uracil, er sammensatt av to nitrogenatomer og fire karbonatomer, og danner en seks-leddet ring. Hvert atom har sitt eget tall fra 1 til 6.
Puriner (adenin og guanin) består av en pyrimidin og en imidazol eller to heterosykler. Purinbasemolekylet er representert av fire nitrogenatomer og fem karbonatomer. Hvert atom er nummerert fra 1 til 9.
Som et resultat av kombinasjonen av en nitrogenholdig base og en pentoserest, dannes et nukleosid. Et nukleotid er en forbindelse av et nukleosid og en fosfatgruppe.
Dannelse av fosfodiesterbindinger
Det er viktig å forstå spørsmålet om hvordan nukleotider er koblet til en polypeptidkjede og danner et nukleinsyremolekyl. Dette skjer på grunn av de såkalte fosfodiesterbindingene.
Samspillet mellom to nukleotider produserer et dinukleotid. Dannelsen av en ny forbindelse skjer ved kondensering, når en fosfodiesterbinding oppstår mellom fosfatresten til en monomer og hydroksygruppen til pentosen til en annen.
Polynukleotidsyntese er en gjentatt repetisjon av denne reaksjonen (flere millioner ganger). Polynukleotidkjeden bygges gjennom dannelsen av fosfodiesterbindinger mellom det tredje og femte karbon av sukker (3C´ og 5C´).
Polynukleotidsammenstilling er en kompleks prosess som skjer med deltakelse av enzymet DNA-polymerase, som sikrer vekst av en kjede fra kun en ende (3´) med en fri hydroksygruppe.
DNA-molekylstruktur
Et DNA-molekyl, som et protein, kan ha en primær, sekundær og tertiær struktur.

Sekvensen av nukleotider i en DNA-kjede bestemmer dens primære; den dannes på grunn av hydrogenbindinger, som er grunnlaget for komplementaritetsprinsippet. Med andre ord, under syntesen av en dobbeltkjede, gjelder et visst mønster: adenin i en kjede tilsvarer tymin i den andre, guanin til cytosin og omvendt. Par av adenin og tymin eller guanin og cytosin dannes på grunn av to i det første og tre i det siste tilfellet hydrogenbindinger. Denne koblingen av nukleotider sikrer en sterk kobling av kjedene og lik avstand mellom dem.
Når du kjenner nukleotidsekvensen til en DNA-streng, kan den andre fullføres ved å bruke prinsippet om komplementaritet eller addisjon.
Den tertiære strukturen til DNA dannes på grunn av komplekse tredimensjonale bindinger, noe som gjør molekylet mer kompakt og i stand til å passe inn i et lite cellevolum. For eksempel er lengden på DNAet til E. coli mer enn 1 mm, mens lengden på cellen er mindre enn 5 mikron.
Antall nukleotider i DNA, nemlig deres kvantitative forhold, følger Chergaff-regelen (antall purinbaser er alltid lik antall pyrimidinbaser). Avstanden mellom nukleotidene er en konstant verdi, lik 0,34 nm, som er deres molekylvekt.
Strukturen til et RNA-molekyl
RNA er representert av en enkelt polynukleotidkjede dannet mellom en pentose (i dette tilfellet ribose) og en fosfatrest. Den er mye kortere enn DNA. Det er også forskjeller i artssammensetningen av nitrogenholdige baser i nukleotidet. I RNA brukes uracil i stedet for pyrimidinbasen tymin. Avhengig av funksjonene som utføres i kroppen, kan RNA være av tre typer.

Ribosomal (rRNA) - inneholder vanligvis fra 3000 til 5000 nukleotider. Etter behov strukturell komponent tar del i dannelsen av det aktive sentrum av ribosomer, stedet for en av de viktigste prosessene i cellen - proteinbiosyntese.
. Transport (tRNA) - består av gjennomsnittlig 75 - 95 nukleotider, utfører overføringen av den ønskede aminosyren til stedet for polypeptidsyntese i ribosomet. Hver type tRNA (minst 40) har sin egen unike sekvens av monomerer eller nukleotider.
. Informasjon (mRNA) - svært variert i nukleotidsammensetning. Overfører genetisk informasjon fra DNA til ribosomer og fungerer som en matrise for syntese av proteinmolekyler.
Rollen til nukleotidene i kroppen
Nukleotider i cellen utfører en rekke viktige funksjoner:
Brukes som byggesteiner for nukleinsyrer (nukleotider i purin- og pyrimidinseriene);
. delta i mange metabolske prosesser i cellen;
. er en del av ATP - den viktigste energikilden i celler;
. fungere som bærere av reduserende ekvivalenter i celler (NAD+, NADP+, FAD, FMN);
. utføre funksjonen til bioregulatorer;
. kan betraktes som andre budbringere for ekstracellulær regelmessig syntese (for eksempel cAMP eller cGMP).
Et nukleotid er en monomer enhet som danner mer komplekse forbindelser - nukleinsyrer, uten hvilke overføring av genetisk informasjon, lagring og reproduksjon er umulig. Frie nukleotider er hovedkomponentene involvert i signal- og energiprosesser som støtter normal funksjon av celler og kroppen som helhet.
I menneskekroppen er det et stort nummer av organiske forbindelser, uten hvilke det er umulig å forestille seg et stabilt forløp av metabolske prosesser som støtter livet til alle. Et av disse stoffene er nukleotider - dette er fosforestere av nukleosider, som spiller en viktig rolle i overføringen av informasjonsdata, samt kjemiske reaksjoner med frigjøring av intracellulær energi.
Som uavhengige organiske enheter danner de fyllingssammensetningen av alle nukleinsyrer og de fleste koenzymer. La oss se nærmere på hva nukleosidfosfater er og hvilken rolle de spiller i menneskekroppen.
Hva består stoffet nukleotid av? Det regnes som en ekstremt ester, som tilhører gruppen av fosforsyrer og nukleosider, som i sine biokjemiske egenskaper er klassifisert som N-glykosider og inneholder heterosykliske fragmenter assosiert med glukosemolekyler og et nitrogenatom.
I naturen er de vanligste nukleotidene DNA.
I tillegg skiller de også organisk materiale med lignende strukturelle egenskaper: ribonukleotider, så vel som deoksyribonukleotider. Alle av dem, uten unntak, er monomere molekyler som tilhører biologiske stoffer av polymertypen som er komplekse i struktur.
Fra dem dannes RNA og DNA fra alle levende vesener, alt fra de enkleste mikroorganismer og virusinfeksjoner til menneskekroppen.
Resten av den molekylære strukturen til fosfor blant nukleosidfosfatene danner en esterbinding med to, tre og i noen tilfeller med fem hydroksylgrupper samtidig. Nesten alle nukleotider, uten unntak, er essensielle stoffer som er dannet fra ortofosforsyrerester, derfor er bindingene deres stabile og desintegrerer ikke under påvirkning av ugunstige faktorer i det indre og ytre miljøet.
Merk! Strukturen til nukleotidene er alltid kompleks og er basert på monoestere. Nukleotidsekvensen kan endres under påvirkning av stressfaktorer.
Biologisk rolle
Påvirkningen av nukleotider på forløpet av alle prosesser i kroppen til levende vesener studeres av forskere som forsker molekylær struktur intracellulært rom.
Basert på laboratoriefunn oppnådd basert på resultatene av mange års arbeid fra forskere fra hele verden, skilles følgende rolle til nukleosidfosfater:
- en universell kilde til vital energi, på grunn av hvilken cellene får næring og følgelig vedlikeholdes normal operasjon vev som danner indre organer, biologiske væsker, epiteldekke og det vaskulære systemet;
- er transportører av glukosemonomerer i celler av enhver type (dette er en av formene for karbohydratmetabolisme, når konsumert sukker, under påvirkning av fordøyelsesenzymer, omdannes til glukose, som transporteres til hvert hjørne av kroppen sammen med nukleosidfosfater );
- utføre funksjonen til et koenzym (vitamin- og mineralforbindelser som bidrar til å gi cellene næringsstoffer);
- komplekse og sykliske mononukleotider er biologiske ledere av hormoner som sprer seg sammen med blodstrømmen, og forsterker også effekten av nevrale impulser;
- allosterisk regulere aktiviteten til fordøyelsesenzymer produsert av bukspyttkjertelvev.
Nukleotider er en del av nukleinsyrer. De er forbundet med tre og fem fosfodiesterbindinger. Genetikere og forskere som har viet livet til molekylærbiologi fortsetter laboratorieforskning på nukleosidfosfater, så hvert år lærer verden enda mer interessante ting om egenskapene til nukleotider.
 Nukleotidsekvensen er en type genetisk likevekt og balanse av arrangementet av aminosyrer i DNA-strukturen, en særegen rekkefølge for plassering av esterrester i sammensetningen av nukleinsyrer.
Nukleotidsekvensen er en type genetisk likevekt og balanse av arrangementet av aminosyrer i DNA-strukturen, en særegen rekkefølge for plassering av esterrester i sammensetningen av nukleinsyrer.
Det bestemmes ved hjelp av den tradisjonelle metoden for sekvensering av biologisk materiale valgt for analyse.
T - tymin;
A – adenin;
G - guanin;
C - cytosin;
R – GA adenin i kompleks med guanin- og purinbaser;
Y – TC pyrimidinforbindelser;
K – GT-nukleotider som inneholder en ketogruppe;
M – AC inkludert i aminogruppen;
S – GC kraftig, kjennetegnet ved tre hydrogenforbindelser;
W – AT er ustabile og danner bare to hydrogenbindinger.
Sekvensen av nukleotider kan endres, og betegnelser med latinske bokstaver er nødvendige i tilfeller der rekkefølgen av arrangement av eterforbindelser er ukjent, er uviktig, eller resultatene av primærforskning allerede eksisterer.
Det største antallet varianter og kombinasjoner av nukleosidfosfater er karakteristisk for DNA. For å skrive RNA-eterforbindelser er det tilstrekkelig med symbolene A, C, G, U. Den siste bokstavbetegnelsen er stoffet uridin, som kun finnes i RNA. En sekvens av symbolske notasjoner skrives alltid uten mellomrom.
Nyttig video: nukleinsyrer (DNA og RNA)
Hvor mange nukleotider er det i DNA
For å forstå så detaljert som mulig hva vi snakker om, bør du ha en klar forståelse av selve DNA. Dette er en egen type molekyler som har en langstrakt form og består av strukturelle elementer, nemlig nukleosidfosfater. Hvor mange nukleotider er det i DNA? Det er 4 typer eterforbindelser av denne typen som er en del av DNA. Disse er adenin, tymin, cytosin og guanin. De danner alle en enkelt kjede, hvorfra den molekylære strukturen til DNA dannes.
Strukturen til DNA ble først dechiffrert tilbake i 1953 av amerikanske forskere Francis Crick og James Watson. Ett molekyl deoksyribonukleinsyre inneholder to kjeder av nukleosidfosfater. De er plassert på en slik måte at de ser ut som en spiral som vrir seg rundt sin akse.
Merk! Antall nukleotider i DNA er konstant og begrenset til bare fire typer - denne oppdagelsen har brakt menneskeheten nærmere å dechiffrere den fullstendige menneskelige genetiske koden.

I dette tilfellet har strukturen til molekylet en viktig funksjon. Alle nukleotidkjeder har egenskapen komplementaritet. Dette betyr at kun eteriske forbindelser av en bestemt type er plassert overfor hverandre. Det er kjent at adenin alltid er plassert overfor tymin. Ingen andre stoffer unntatt guanin kan finnes på motsatt side av cytosin. Slike nukleotidpar danner komplementaritetsprinsippet og er uadskillelige.
Vekt og lengde
 Ved hjelp av komplekse matematiske beregninger og laboratoriestudier klarte forskerne å fastslå de eksakte fysiske og biologiske egenskapene til eterforbindelsene som danner den molekylære strukturen til deoksyribonukleinsyre.
Ved hjelp av komplekse matematiske beregninger og laboratoriestudier klarte forskerne å fastslå de eksakte fysiske og biologiske egenskapene til eterforbindelsene som danner den molekylære strukturen til deoksyribonukleinsyre.
Det er kjent at den utvidede lengden av en intracellulær rest bestående av aminosyrer i en enkelt polypeptidkjede er 3,5 ångstrøm. Gjennomsnittlig masse av en molekylrest er 110 amu.
I tillegg isoleres også monomerer av nukleotidtypen, som er dannet ikke bare fra aminosyrer, men også har eterkomponenter. Dette er monomerer av DNA og RNA. Deres lineære lengde måles direkte inne i nukleinsyren og er minst 3,4 ångstrøm. Molekylvekten til ett nukleosidfosfat er innenfor 345 amu. Dette er de første dataene som brukes i praksis laboratoriearbeid dedikert til eksperimenter, genetisk forskning og andre vitenskapelige aktiviteter.
Medisinske betegnelser
Genetikk, som vitenskap, utviklet seg tilbake i perioden da det ikke var forskning på strukturen til DNA i mennesker og andre levende vesener. molekylært nivå. Derfor, i løpet av perioden med pre-molekylær genetikk, ble nukleotidbindinger utpekt som det minste elementet i strukturen til DNA-molekylet. Både tidligere og for tiden har essensielle stoffer av denne typen vært utsatt for. Det kan være spontant eller indusert, og det er grunnen til at begrepet "recon" også brukes for å betegne nukleosidfosfater med en skadet struktur.
For å definere konseptet for forekomsten av en mulig mutasjon i nitrogenholdige forbindelser av nukleotidbindinger, brukes begrepet "mouton". Disse betegnelsene er mer etterspurt i laboratoriearbeid med biologisk materiale. De brukes også av genetiske forskere som studerer strukturen til DNA-molekyler, måtene å overføre arvelig informasjon på, metoder for å kode den og mulige kombinasjoner av gener oppnådd som et resultat av sammensmeltingen av det genetiske potensialet til to seksuelle partnere.
I kontakt med
 Great Tartary - the Rus Empire Om menneskehetens eldgamle historie
Great Tartary - the Rus Empire Om menneskehetens eldgamle historie Prins Ivan Vasilyevich 1472
Prins Ivan Vasilyevich 1472 "Historien om A.P. Platonov "Den ukjente blomsten" (minianmeldelse). Om Andrei Platonovs historie "Den ukjente blomsten" Hva det sanne eventyret Den ukjente blomsten lærte meg
"Historien om A.P. Platonov "Den ukjente blomsten" (minianmeldelse). Om Andrei Platonovs historie "Den ukjente blomsten" Hva det sanne eventyret Den ukjente blomsten lærte meg