Problemet med lokalisering av funksjoner i hjernebarken. Lokalisering av funksjoner i hjernebarken
Hjernebarken er den evolusjonsmessig yngste formasjonen, som hos mennesker har nådd sine største verdier i forhold til resten av hjernemassen. Hos mennesker er massen av hjernebarken i gjennomsnitt 78 % av hjernens totale masse. Hjernebarken er ekstremt viktig i reguleringen av kroppens vitale funksjoner, implementeringen av komplekse former for atferd og utviklingen av nevropsykiske funksjoner. Disse funksjonene leveres ikke bare av hele massen av det kortikale stoffet, men også av de ubegrensede mulighetene for assosiative forbindelser mellom cellene i cortex og subkortikale formasjoner, noe som skaper forhold for den mest komplekse analysen og syntesen av innkommende informasjon, for utvikling av læringsformer som er utilgjengelige for dyr.
Når vi snakker om den ledende rollen til hjernebarken i nevrofysiologiske prosesser, bør vi ikke glemme at denne høyere avdelingen kan fungere normalt bare i nært samspill med subkortikale formasjoner. Kontrasten mellom cortex og underliggende deler av hjernen er i stor grad skjematisk og betinget. I i fjor ideer om den vertikale organiseringen av funksjonene til nervesystemet og om sirkulære kortikale-subkortikale forbindelser utvikles.
Cellene i cortex er spesialiserte i mye mindre grad enn kjernene i de subkortikale formasjonene. Det følger at de kompenserende evnene til cortex er svært høye - funksjonene til de berørte cellene kan overtas av andre nevroner; skader på ganske store områder av cortex kan klinisk virke svært uskarpe (de såkalte kliniske stille soner). Fraværet av smal spesialisering av kortikale nevroner skaper forhold for fremveksten av et bredt utvalg av interneuronforbindelser, dannelsen av komplekse "ensembler" av nevroner som regulerer forskjellige funksjoner. Dette er det viktigste grunnlaget for læringsevne. Det teoretisk mulige antallet forbindelser mellom de 14 milliarder cellene i hjernebarken er så stort at i løpet av en persons liv forblir en betydelig del av dem ubrukt. Dette bekrefter nok en gang de ubegrensede mulighetene for menneskelig læring.
Til tross for den kjente uspesifisiteten til kortikale celler, er visse grupper av dem anatomisk og funksjonelt nærmere knyttet til visse spesialiserte deler av nervesystemet. Den morfologiske og funksjonelle tvetydigheten til forskjellige områder av cortex lar oss snakke om de kortikale sentrene for syn, hørsel, berøring, etc., som har en spesifikk lokalisering. I verkene til forskere på 1800-tallet ble dette lokaliseringsprinsippet tatt til det ytterste: det ble gjort forsøk på å identifisere sentre for vilje, tenkning, evnen til å forstå kunst osv. For tiden ville det være feil å snakke om kortikalt senter som en strengt begrenset gruppe celler. Det skal bemerkes at spesialiseringen av nervekoblinger dannes i løpet av livet.
I følge I.P. Pavlov består hjernesenteret, eller den kortikale delen av analysatoren, av en "kjerne" og "spredte elementer." "Kjernen" er en relativt morfologisk homogen gruppe av celler med en presis projeksjon av reseptorfelt. "Sprede elementer" er plassert i en sirkel eller i en viss avstand fra "kjernen": de utfører en mer elementær og mindre differensiert analyse og syntese av innkommende informasjon.
Av de 6 lagene med kortikale celler er de øvre lagene mest utviklet hos mennesker sammenlignet med lignende lag hos dyr og dannes i ontogenese mye senere enn de nedre lagene. De nedre lagene av cortex har forbindelser med perifere reseptorer (lag IV) og med muskler (lag V) og kalles "primære" eller "projeksjon" kortikale soner på grunn av deres direkte forbindelse med de perifere delene av analysatoren. Over de "primære" sonene er bygget systemer av "sekundære" soner (lag II og III), der assosiative forbindelser med andre deler av cortex dominerer, derfor kalles de også projeksjonsassosiative.
Dermed identifiseres to grupper av cellulære soner i de kortikale representasjonene til analysatorene. En slik struktur finnes i den oksipitale sonen, der synsbanene projiseres, i den tidsmessige sonen, hvor de auditive banene slutter, i den bakre sentrale gyrus - den kortikale delen av den sensitive analysatoren, i den fremre sentrale gyrusen - den kortikale motorsenter. Den anatomiske heterogeniteten til de "primære" og "sekundære" sonene er ledsaget av fysiologiske forskjeller. Eksperimenter med stimulering av cortex har vist at stimulering av de primære sonene i sensoriske områder fører til fremveksten av elementære sensasjoner. For eksempel forårsaker irritasjon av oksipitale områder en følelse av flimrende lyspunkter, linjer osv. Ved irritasjon av sekundærsonene oppstår mer komplekse fenomener: subjektet ser forskjellig utformede objekter - mennesker, fugler osv. Det kan antas at det er i sekundærsonene operasjoner utføres gnosis og delvis praxis.
I tillegg skilles tertiære soner, eller soner med overlapping av kortikale representasjoner av individuelle analysatorer, i cortex. Hos mennesker okkuperer de et veldig betydelig sted og befinner seg først og fremst i parieto-temporo-occipital regionen og i frontalsonen. Tertiære soner inngår omfattende forbindelser med kortikale analysatorer og sikrer dermed utviklingen av komplekse, integrerende reaksjoner, blant hvilke meningsfulle handlinger inntar førsteplassen hos mennesker. I de tertiære sonene foregår derfor planleggings- og kontrolloperasjoner, som krever kompleks deltakelse fra forskjellige deler av hjernen.
I tidlig barndom overlapper de funksjonelle sonene i cortex hverandre, deres grenser er diffuse, og bare i prosessen praktiske aktiviteter Det er en konstant konsentrasjon av funksjonelle soner i avgrensede sentre atskilt fra hverandre. I klinikken opplever voksne pasienter svært konstante symptomkomplekser når visse områder av cortex og tilhørende nervebaner påvirkes.
I barndommen, på grunn av ufullstendig differensiering av funksjonelle soner, kan fokal skade på hjernebarken ikke ha en klar klinisk manifestasjon, som bør huskes når man vurderer alvorlighetsgraden og grensene for hjerneskade hos barn.
I funksjonelle termer kan vi skille de viktigste integrerende nivåene av kortikal aktivitet.
Det første signalsystemet er assosiert med aktivitetene til individuelle analysatorer og utfører de primære stadiene av gnosis og praksis, dvs. integrering av signaler som kommer gjennom kanalene til individuelle analysatorer, og dannelsen av responshandlinger som tar hensyn til tilstanden til eksterne og Internt miljø, samt tidligere erfaringer. Dette første nivået inkluderer visuell persepsjon av objekter med konsentrasjon av oppmerksomhet på enkelte av dets detaljer, frivillige bevegelser med aktiv styrking eller hemming av dem.
Et mer komplekst funksjonelt nivå av kortikal aktivitet forener systemene til forskjellige analysatorer, inkluderer et andre signalsystem), forener systemene til forskjellige analysatorer, noe som gjør det mulig for en meningsfull oppfatning av miljøet, en holdning til omverdenen "med kunnskap og forståelse.» Dette integreringsnivået er nært knyttet til taleaktivitet, og forståelsen av tale (talegnosis) og bruken av tale som et middel for adressering og tenkning (talepraxis) er ikke bare sammenkoblet, men også bestemt av ulike nevrofysiologiske mekanismer, som er av stor klinisk betydning.
Det høyeste nivået av integrering dannes i en person i ferd med å modnes som et sosialt vesen, i ferd med å mestre ferdighetene og kunnskapen som samfunnet har.
Det tredje stadiet av kortikal aktivitet spiller rollen som en slags avsender av komplekse prosesser med høyere nervøs aktivitet. Det sikrer hensiktsmessigheten til visse handlinger, og skaper betingelser for best mulig gjennomføring. Dette oppnås ved å "filtrere" signaler som for tiden er av størst betydning fra signaler av sekundær betydning, utføre sannsynlige prognoser for fremtiden og danne langsiktige oppgaver.
Selvfølgelig kunne kompleks kortikal aktivitet ikke utføres uten deltakelse fra informasjonslagringssystemet. Derfor er minnemekanismer en av de viktigste komponentene i denne aktiviteten. I disse mekanismene flyter ikke bare funksjonene til å registrere informasjon (memorisering), men også funksjonene for å skaffe nødvendig informasjon fra "minne" (minne), samt funksjonene til å overføre informasjon fra blokker med RAM (det som trengs for øyeblikket) inn i langtidsminneblokker og omvendt. Ellers ville det være umulig å lære nye ting, siden gamle ferdigheter og kunnskaper ville forstyrre dette.
Nyere nevrofysiologiske studier har gjort det mulig å fastslå hvilke funksjoner som er overveiende karakteristiske for visse deler av hjernebarken. Selv i forrige århundre var det kjent at den oksipitale regionen av cortex er nært forbundet med den visuelle analysatoren, den temporale regionen - med den auditive (Heschls gyrus), smaksanalysatoren, den fremre sentrale gyrusen - med motoren, og bakre sentrale gyrus - med den muskulokutane analysatoren. Vi kan betinget anta at disse avdelingene er assosiert med den første typen kortikal aktivitet og gir de enkleste formene for gnosis og praxis.
Deler av cortex lokalisert i parietotemporal-occipital regionen tar en aktiv del i dannelsen av mer komplekse gnostisk-praksiske funksjoner. Skader på disse områdene fører til mer komplekse former for lidelser. Wernickes gnostiske talesenter ligger i tinninglappen på venstre hjernehalvdel. Det motoriske talesenteret er lokalisert noe foran den nedre tredjedelen av den fremre sentrale gyrusen (Brocas sentrum). I tillegg til sentrene for muntlig tale, er det sensoriske og motoriske sentre for skriftlig tale og en rekke andre formasjoner, på en eller annen måte knyttet til tale. Den parieto-temporo-occipitale regionen, der banene som kommer fra ulike analysatorer lukkes, er av største betydning for dannelsen av høyere mentale funksjoner. Den kjente nevrofysiologen og nevrokirurgen W. Penfield kalte dette området den tolkende cortex. I dette området er det også formasjoner involvert i minnemekanismer.
Spesiell vekt er lagt til frontalregionen. I følge moderne konsepter er det denne delen av hjernebarken som tar en aktiv del i organiseringen av målrettet aktivitet, i langsiktig planlegging og bestemmelse, det vil si at den tilhører den tredje typen kortikale funksjoner.
Hovedsentrene i hjernebarken. Frontallappen. Motoranalysatoren er plassert i fremre sentrale gyrus og parasentral lobule (Brodmanns områder 4, 6 og 6a). I mellomlagene er det en analysator av kinestetiske stimuli som kommer fra skjelettmuskulatur, sener, ledd og bein. I lag V og delvis VI er gigantiske pyramidale celler av Betz lokalisert, hvis fibre danner pyramidebanen. Den fremre sentrale gyrus har en viss somatotopisk projeksjon og er forbundet med den motsatte halvdelen av kroppen. Musklene i underekstremitetene projiseres i de øvre delene av gyrusen, og musklene i ansiktet i de nedre delene. Stammen, strupehodet og svelget er representert i begge halvkuler (fig. 55).
Rotasjonssenteret for øynene og hodet i motsatt retning er plassert i den midtre frontale gyrusen i det premotoriske området (felt 8, 9). Arbeidet til dette senteret er nært forbundet med systemet til den bakre langsgående fasciculus, de vestibulære kjernene, formasjoner av det striopallidale systemet, som er involvert i reguleringen av torsjon, så vel som med den kortikale delen av den visuelle analysatoren (felt 17) ).
I de bakre delene av den øvre frontale gyrus er det et senter som gir opphav til fronto-pontocerebellarbanen (felt 8). Dette området av hjernebarken er involvert i å sikre koordinering av bevegelser knyttet til oppreist holdning, opprettholde balanse mens du står og sitter, og regulerer arbeidet til den motsatte hjernehalvdelen av lillehjernen.
Det motoriske talesenteret (speech praxis center) ligger i den bakre delen av den nedre frontale gyrus - Brocas gyrus (område 44). Senteret gir analyse av kinestetiske impulser fra musklene i det talemotoriske apparatet, lagring og implementering av "bilder" av taleautomatikk, dannelsen av muntlig tale, og er nært forbundet med plasseringen bak den av den nedre delen av den. fremre sentrale gyrus (projeksjonssone av leppene, tungen og strupehodet) og med den som ligger foran det musikalsk motorisk senter.
Det musikalske motoriske senteret (felt 45) gir en viss tonalitet, modulering av tale, samt evne til å komponere musikalske fraser og synge.
Sentrum av skriftlig tale er lokalisert i den bakre delen av den midtre frontale gyrusen i umiddelbar nærhet til det kortikale projeksjonsområdet på hånden (felt 6). Senteret sørger for automatikk i skrivingen og henger funksjonelt sammen med Brocas senter.
Parietallapp. Sentrum av hudanalysatoren er plassert i den bakre sentrale gyrusen til felt 1, 2, 3 og cortex i den øvre parietale regionen (felt 5 og 7). I den bakre sentrale gyrus projiseres taktil, smerte og temperaturfølsomhet i den motsatte halvdelen av kroppen. Følsomheten til benet projiseres i de øvre delene, og følsomheten til ansiktet projiseres i de nedre delene. Boks 5 og 7 representerer elementer av dyp følsomhet. Posteriort til de midtre delene av den bakre sentrale gyrusen er senteret for stereognose (felt 7,40 og delvis 39), som gir muligheten til å gjenkjenne objekter ved berøring.
Posteriort til de øvre delene av den bakre sentrale gyrusen er et senter som gir evnen til å gjenkjenne sin egen kropp, dens deler, deres proporsjoner og relative posisjoner (felt 7).
Sentrum av praxis er lokalisert i den nedre parietale lobulen til venstre, den supramarginale gyrus (felt 40 og 39). Senteret sørger for lagring og implementering av bilder av motorautomater (praxisfunksjoner).
I de nedre delene av den fremre og bakre sentrale gyri er det sentrum av analysatoren for interoceptive impulser av indre organer og blodkar. Senteret har nære forbindelser med subkortikale vegetative formasjoner.
Tinninglappen. Sentrum av den auditive analysatoren er plassert i den midtre delen av den øvre temporal gyrus, på overflaten som vender mot insulaen (Heschls gyrus, områder 41, 42, 52). Disse formasjonene gir projeksjon av sneglehuset, samt lagring og gjenkjennelse av auditive bilder.
Sentrum av den vestibulære analysatoren (felt 20 og 21) er plassert i de nedre delene av den ytre overflaten av tinninglappen, er projeksjon, og er i nær forbindelse med de nedre basale delene av tinninglappene, og gir opphav til den occipitotemporale cortical-pontine-cerebellar bane.
Ris. 55. Opplegg for lokalisering av funksjoner i hjernebarken (A - D). I - projeksjonsmotorsone; II - sentrum for rotasjon av øyne og hode i motsatt retning; III - projeksjonsfølsomhetssone; IV - projeksjon visuell sone; projeksjon gnostiske soner: V - hørsel; VI - lukt, VII - smak, VIII - gnostisk sone av kroppsdiagrammet; IX - stereognose sone; X - gnostisk visuell sone; XI - Gnostisk lesesone; XII - gnostisk talesone; XIII - praksissone; XIV - praktisk talesone; XV - praktisk skrivesone; XVI - sone for kontroll over funksjonen til lillehjernen.
Sentrum av lukteanalysatoren er plassert i den fylogenetisk eldste delen av hjernebarken - i kroken og ammonhornet (felt 11a, e) og gir projeksjonsfunksjon, samt lagring og gjenkjenning av luktbilder.
Sentrum av smaksanalysatoren er plassert i umiddelbar nærhet av sentrum av luktanalysatoren, dvs. i kroken og ammonhornet, men i tillegg i den nederste delen av den bakre sentrale gyrusen (område 43), samt i øya. I likhet med luktanalysatoren gir senteret projeksjonsfunksjon, lagring og gjenkjenning av smaksbilder.
Det akustisk-gnostiske sansetalesenteret (Wernickes sentrum) er lokalisert i de bakre delene av den øvre temporal gyrus til venstre, i dybden av lateral sulcus (område 42, samt områder 22 og 37). Senteret gir gjenkjenning og lagring av lydbilder av muntlig tale, både egne og andres.
I umiddelbar nærhet av Wernickes sentrum (den midterste tredjedelen av den overordnede temporale gyrus - område 22) er det et senter som sikrer gjenkjennelse av musikalske lyder og melodier.
Bakhode lapp. Sentrum av den visuelle analysatoren er plassert i occipitallappen (felt 17, 18, 19). Felt 17 er en visuell projeksjonssone, felt 18 og 19 gir lagring og gjenkjenning av visuelle bilder, visuell orientering i et uvanlig miljø.
På grensen til tinninglappen, occipitallappen og parietallappen er senteret til den skriftlige taleanalysatoren (felt 39), som er nært forbundet med Wernicke-senteret i tinninglappen, med midten av den visuelle analysatoren til occipitallappen, som samt med sentrene av parietallappen. Lesesenteret gir gjenkjenning og lagring av skriftspråklige bilder.
Data om lokalisering av funksjoner ble oppnådd enten som et resultat av irritasjon av ulike deler av cortex i et eksperiment, eller som et resultat av analyse av forstyrrelser som oppsto som følge av skade på visse områder av cortex. Begge disse tilnærmingene kan bare indikere deltakelsen av visse kortikale soner i visse mekanismer, men betyr ikke i det hele tatt deres strenge spesialisering eller entydig forbindelse med strengt definerte funksjoner.
I den nevrologiske klinikken, i tillegg til tegn på skade på områder av hjernebarken, er det symptomer på irritasjon av dens individuelle områder. I tillegg, i barndommen, observeres fenomener med forsinket eller svekket utvikling av kortikale funksjoner, noe som betydelig endrer de "klassiske" symptomene. Eksistensen av forskjellige funksjonelle typer kortikal aktivitet forårsaker forskjellige symptomer på kortikale lesjoner. Analyse av disse symptomene lar oss identifisere arten av lesjonen og dens plassering.
Avhengig av type kortikal aktivitet, er det mulig å skille mellom kortikale lesjoner forstyrrelser av gnosis og praksis på ulike nivåer integrering; taleforstyrrelser på grunn av deres praktiske betydning; forstyrrelser i regulering av målrettethet, målrettethet av nevrofysiologiske funksjoner. Med hver type lidelse kan minnemekanismene som er involvert i et gitt funksjonssystem også bli forstyrret. I tillegg er mer total hukommelsessvikt mulig. I tillegg til relativt lokale kortikale symptomer, observeres også mer diffuse symptomer i klinikken, som først og fremst viser seg ved utviklingshemming og atferdsforstyrrelser. Begge disse lidelsene er av spesiell betydning i barnepsykiatrien, selv om i hovedsak mange varianter av slike lidelser kan betraktes som grense mellom nevrologi, psykiatri og pediatri.
Studiet av kortikale funksjoner i barndommen har en rekke forskjeller fra studiet av andre deler av nervesystemet. Det er viktig å etablere kontakt med barnet og opprettholde en avslappet omgangstone med det. Siden mange diagnostiske oppgaver presentert for et barn er svært komplekse, må man strebe etter å sikre at han ikke bare forstår oppgaven, men også blir interessert i den. Noen ganger når man undersøker barn som er altfor distraherte, motorisk dehemmede eller psykisk utviklingshemmede, må man bruke mye tålmodighet og oppfinnsomhet for å identifisere eksisterende abnormiteter. I mange tilfeller blir analysen av et barns kortikale funksjoner hjulpet av foreldrenes rapporter om hans oppførsel hjemme, på skolen og skolens egenskaper.
Når man studerer kortikale funksjoner er det viktig psykologisk eksperiment, hvor essensen er presentasjonen av standardiserte, målrettede oppgaver. Visse psykologiske metoder lar en vurdere visse aspekter ved mental aktivitet isolert, mens andre lar dem vurderes mer omfattende. Disse inkluderer såkalte personlighetstester.
Gnosis og dens lidelser. Gnosis betyr bokstavelig talt anerkjennelse. Vår orientering i omverdenen er assosiert med å gjenkjenne objekters form, størrelse, romlige forhold og til slutt å forstå deres betydning, som ligger i objektets navn. Denne beholdningen av informasjon om verden rundt består av analyse og syntese av sensoriske impulsstrømmer og lagres i minnesystemer. Reseptorapparatet og overføringen av sensoriske impulser med lesjoner av høyere gnostiske mekanismer er bevart, men tolkningen av disse impulsene og sammenligningen av mottatte data med bilder lagret i minnet blir forstyrret. Som et resultat oppstår en forstyrrelse av gnosis - agnosia, hvis essens er at mens oppfatningen av objekter er bevart, går følelsen av deres "kjenthet" tapt og verden, tidligere så kjent i detaljer, blir fremmed, uforståelig, blottet for mening.
Men gnosis kan ikke tenkes som en enkel sammenligning, gjenkjennelse av et bilde. Gnosis er en prosess med kontinuerlig oppdatering, avklaring, konkretisering av bildet som er lagret i minnematrisen, under påvirkning av dens gjentatte sammenligning med den mottatte informasjonen.
Total agnosi, hvor fullstendig desorientering observeres, er sjelden. Mye oftere blir gnose forstyrret i ethvert analytisk system, og avhengig av graden av skade varierer alvorlighetsgraden av agnosia.
Visuell agnosi oppstår når occipital cortex er skadet. Pasienten ser gjenstanden, men kjenner den ikke igjen. Det kan være ulike alternativer her. I noen tilfeller beskriver pasienten riktig de ytre egenskapene til et objekt (farge, form, størrelse), men kan ikke gjenkjenne objektet. For eksempel beskriver en pasient et eple som "noe rundt og rosa", uten å gjenkjenne eplet som et eple. Men hvis du gir denne gjenstanden til pasienten, vil han gjenkjenne den når han kjenner den. Det er tider når pasienten ikke kjenner igjen kjente ansikter. Noen pasienter med en lignende lidelse blir tvunget til å huske mennesker basert på andre egenskaper (klær, føflekk osv.). I andre tilfeller av agnosi gjenkjenner pasienten et objekt, navngir dets egenskaper og funksjon, men kan ikke huske hva det heter. Disse tilfellene tilhører gruppen taleforstyrrelser.
Ved noen former for visuell agnosi er romlig orientering og visuell hukommelse svekket. I praksis, selv om et objekt ikke gjenkjennes, kan vi snakke om brudd på minnemekanismer, siden det oppfattede objektet ikke kan sammenlignes med bildet i den gnostiske matrisen. Men det er også tilfeller der, når en gjenstand presenteres igjen, sier pasienten at han allerede har sett den, selv om han fortsatt ikke kan gjenkjenne den. Hvis romlig orientering er svekket, gjenkjenner ikke pasienten bare tidligere kjente ansikter, hus osv., men kan også gå på samme sted mange ganger uten å vite det.
Ofte, med visuell agnosi, lider også gjenkjenningen av bokstaver og tall, og det oppstår tap av leseevne. Den isolerte typen av denne lidelsen vil bli analysert i analysen av talefunksjon.
For å studere visuell gnosis brukes et sett med objekter. Når de presenterer dem for faget, blir de bedt om å identifisere og beskrive dem. utseende, sammenligne hvilke objekter som er større og hvilke som er mindre. De bruker også et sett med bilder, farger, vanlig og kontur. De evaluerer ikke bare gjenkjennelsen av objekter, ansikter, men også plott. Samtidig kan du teste visuelt minne: presenter flere bilder, bland dem med tidligere usett og be barnet velge kjente bilder. Samtidig er arbeidstid, utholdenhet og tretthet også tatt i betraktning.
Det bør huskes at barn gjenkjenner konturbilder dårligere enn fargede og monokromatiske. Å forstå handlingen er relatert til barnets alder og grad av mental utvikling. Samtidig er agnosi i klassisk form sjelden hos barn på grunn av ufullstendig differensiering av kortikale sentre.
Auditiv agnosi. De oppstår når tinninglappen er skadet i området av Heschls gyrus. Pasienten kan ikke gjenkjenne tidligere kjente lyder: tikken av en klokke, ringing av en bjelle, lyden av rennende vann. Mulig svekkelse av gjenkjennelse av musikalske melodier - amusia. I noen tilfeller blir bestemmelsen av lydretningen forstyrret. Ved noen typer auditiv agnosi er ikke pasienten i stand til å skille frekvensen av lyder, for eksempel metronomslag.
Sensitiv agnosi er forårsaket av svekket gjenkjennelse av taktil, smerte, temperatur, proprioseptive bilder eller deres kombinasjoner. De oppstår når parietalregionen er skadet. Dette inkluderer astereognosis, kroppsdiagramforstyrrelser. I noen varianter av astereognosis kan pasienten ikke bare identifisere et objekt ved berøring, men er heller ikke i stand til å bestemme formen på objektet eller egenskapene til overflaten. Sensitiv agnosi inkluderer også anosognosi, der pasienten ikke er klar over sin defekt, for eksempel lammelse. Fantomfornemmelser kan tilskrives forstyrrelser av sensitiv gnosis.
Når man undersøker barn, bør man huske på at et lite barn ikke alltid kan vise deler av kroppen sin riktig; Det samme gjelder pasienter som lider av demens. I slike tilfeller er det selvfølgelig ikke nødvendig å snakke om en forstyrrelse av kroppsdiagrammet.
Smak og luktagnosi er sjeldne. I tillegg er gjenkjenning av lukt svært individuelt og i stor grad knyttet til personlig erfaring person.
Praxis og dens lidelser. Praxis refererer til målrettet handling. En person lærer mange spesielle motoriske handlinger i løpet av livet. Mange av disse ferdighetene, som dannes med deltagelse av høyere kortikale mekanismer, er automatisert og blir den samme integrerte menneskelige evnen som enkle bevegelser. Men når de kortikale mekanismene som er involvert i gjennomføringen av disse handlingene er skadet, oppstår særegne bevegelsesforstyrrelser - apraksi, der det ikke er noen lammelse, ingen forstyrrelser av tone eller koordinasjon, og til og med enkle frivillige bevegelser er mulige, men mer komplekse, rent menneskelige motoriske handlinger forstyrres. Pasienten opplever plutselig at han ikke er i stand til å utføre slike tilsynelatende enkle handlinger som å håndhilse, feste knapper, gre håret, tenne en fyrstikk osv. Apraksi oppstår først og fremst når den parieto-temporo-occipitale regionen av den dominerende halvkulen er påvirket. I dette tilfellet påvirkes begge halvdelene av kroppen. Apraksi kan også oppstå ved skade på den subdominante høyre hemisfæren (hos høyrehendte) og corpus callosum, som forbinder begge hjernehalvdelene. I dette tilfellet oppdages apraksi bare til venstre. Med apraksi lider handlingsplanen, det vil si dannelsen av en kontinuerlig kjede av motorautomater. Her er det på sin plass å sitere ordene til K. Marx: «Menneskelig handling skiller seg fra arbeidet til «den beste bien» ved at en person allerede har bygget i hodet før han bygger. På slutten av arbeidsprosessen oppnås et resultat som allerede var ideelt før starten av denne prosessen, det vil si i arbeiderens sinn.»
På grunn av brudd på handlingsplanen, når han prøver å fullføre en oppgave, gjør pasienten mange unødvendige bevegelser. I noen tilfeller observeres parapraksi når en handling utføres som bare vagt minner om den gitte oppgaven. Noen ganger observeres også utholdenhet, det vil si å bli sittende fast på noen handlinger. For eksempel blir pasienten bedt om å gjøre en innbydende bevegelse med hånden. Etter å ha fullført denne oppgaven tilbyr de å logre med fingeren, men pasienten utfører fortsatt den første handlingen.
I noen tilfeller, med apraksi, blir vanlige, hverdagslige handlinger bevart, men profesjonelle ferdigheter går tapt (for eksempel evnen til å bruke et fly, skrutrekker, etc.).
I henhold til kliniske manifestasjoner skilles flere typer apraksi ut: motorisk, ideell og konstruktiv.
Motorisk apraksi. Pasienten kan ikke utføre handlinger i henhold til instruksjoner eller til og med etterligning. Han blir bedt om å klippe papir med saks, snøre en sko, streke papir med blyant og linjal osv., men pasienten, selv om han forstår oppgaven, kan ikke fullføre den, og viser fullstendig hjelpeløshet. Selv om du viser hvordan dette gjøres, kan pasienten fortsatt ikke gjenta bevegelsen. I noen tilfeller viser det seg å være umulig å utføre så enkle handlinger som å sitte på huk, snu, klappe i hendene.
Ideatisk apraksi. Pasienten kan ikke utføre handlinger på en oppgave med ekte og imaginære gjenstander (for eksempel vise hvordan man gre håret, røre sukker i et glass osv.), samtidig som handlingene til imitasjon bevares. I noen tilfeller kan pasienten automatisk utføre visse handlinger uten å tenke. For eksempel kan han målrettet ikke feste en knapp, men utfører denne handlingen automatisk.
Konstruktiv apraksi. Pasienten kan utføre ulike handlinger ved imitasjon og ved verbale ordre, men er ikke i stand til å lage en kvalitativ ny motorisk handling, sette sammen en helhet av deler, for eksempel lage en viss figur ut av fyrstikker, sette sammen en pyramide osv.
Noen varianter av apraksi er assosiert med nedsatt gnose. Pasienten kjenner ikke igjen objektet eller kroppsdiagrammet hans er forstyrret, så han klarer ikke å utføre oppgaver eller utfører dem usikkert og ikke helt korrekt.
For å studere praksis tilbys en rekke oppgaver (sett deg ned, rist en finger, gre håret osv.). De blir også presentert for oppgaver for handlinger med imaginære gjenstander (de blir bedt om å vise hvordan de spiser, hvordan de ringer, hvordan de hogger ved osv.). Vurder hvordan pasienten kan imitere handlingene som vises.
Spesielle psykologiske teknikker brukes også for å studere gnosis og praksis. Blant dem er et viktig sted okkupert av Seguin-brett med utsparinger i forskjellige former, der du må sette inn figurer som tilsvarer utsparingene. Denne metoden lar deg også vurdere graden av mental utvikling. Koss-teknikken brukes også: et sett med kuber i forskjellige farger. Fra disse kubene må du sette sammen et mønster som matcher det som vises på bildet. Eldre barn får også tilbud om en Link-kube: de må brette en kube av 27 forskjellige terninger slik at alle sidene har samme farge. Pasienten får vist den sammensatte kuben, så ødelegger de den og ber ham sette den sammen igjen.
I disse teknikkene veldig viktig har hvordan barnet utfører oppgaven: om det handler ved prøving og feiling eller etter en bestemt plan.
Ris. 56. Diagram over koblinger av talesentre og regulering av taleaktivitet.
1 - skrivesenter; 2 - Brocas sentrum; 3 - sentrum av praksis; 4 - sentrum av proprioseptiv gnose; 5 - lesesenter; 6 - Wernicke sentrum; 7 - senter for auditiv gnosis; 8 - sentrum for visuell gnosis.
Det er viktig å huske at praksis utvikler seg etter hvert som barnet modnes, så små barn kan ennå ikke utføre så enkle handlinger som å gre håret, feste knapper osv. Apraxia i sin klassiske form, som agnosia, forekommer hovedsakelig hos voksne.
Tale og dens forstyrrelser. I Visuelle, auditive, motoriske og kinestetiske analysatorer deltar i implementeringen av talefunksjoner, samt skriving og lesing. Av stor betydning er bevaringen av innerveringen av musklene i tungen, strupehodet, den myke ganen, tilstanden til paranasale bihuler og munnhulen, som spiller rollen som resonatorhuler. I tillegg er koordinering av pust og uttale av lyder viktig.
For normal taleaktivitet er koordinert funksjon av hele hjernen og andre deler av nervesystemet nødvendig. Talemekanismer har en kompleks og flertrinns organisasjon (fig. 56).
Tale er den viktigste menneskelige funksjonen, derfor deltar kortikale talesoner lokalisert i den dominerende halvkulen (Brocas og Wernickes sentre), motoriske, kinetiske, auditive og visuelle områder, samt afferente og efferente veier relatert til de pyramidale og ekstrapyramidale systemene. i sin implementering. , analysatorer av følsomhet, hørsel, syn, bulbære deler av hjernen, visuelle, oculomotoriske, ansikts-, auditive-, glossopharyngeale, vagus- og hypoglossale nerver.
Kompleksiteten og flertrinnsnaturen til talemekanismer bestemmer også mangfoldet av taleforstyrrelser. Når innerveringen av taleapparatet blir forstyrret, dysartri- Artikulasjonsforstyrrelse, som kan være forårsaket av sentral eller perifer lammelse av det talemotoriske apparatet, skade på lillehjernen eller det striopallidale systemet.
Det er også dyslalia- fonetisk feil uttale individuelle lyder. Dyslalia kan være funksjonell i naturen og kan ganske vellykket elimineres med logopedøkter. Under alalia forstå forsinkelsen taleutvikling. Vanligvis til V.A. I en alder av 10 år begynner barnet å snakke, men noen ganger skjer dette mye senere, selv om barnet forstår tale rettet til ham godt. Forsinket taleutvikling påvirker også mental utvikling, siden tale er det viktigste informasjonsmidlet for et barn. Imidlertid er det også tilfeller av alalia forbundet med demens. Barnet henger etter i mental utvikling, og derfor dannes ikke talen hans. Disse forskjellige tilfellene av alalia må differensieres, da de har forskjellige prognoser.
Med utviklingen av talefunksjon i den dominerende halvkulen (til venstre for høyrehendte, til høyre for venstrehendte), dannes gnostiske og praktiske talesentre, og deretter - skrive- og lesesentre.
Kortikale taleforstyrrelser er varianter av agnosi og apraksi. Det er uttrykksfull (motorisk) og imponerende (sanselig) tale. Kortikal motorisk taleforstyrrelse er apraxia av tale, sansetale - taleagnosi. I noen tilfeller er tilbakekallingen av de nødvendige ordene svekket, det vil si at minnemekanismer lider. Taleagnosi og apraksi kalles afasi.
Det bør huskes at taleforstyrrelser kan være en konsekvens av generell apraksi (apraksi av stammen, lemmer) eller oral apraksi, der pasienten mister evnen til å åpne munnen, puste ut kinnene og stikke ut tungen. Disse tilfellene er ikke afasi; taleapraksi oppstår her sekundært som en manifestasjon av generelle praksiske lidelser.
Taleforstyrrelser i barndommen, avhengig av årsakene til deres forekomst, kan deles inn i følgende grupper:
I. Taleforstyrrelser forbundet med organisk skade på sentralnervesystemet. Avhengig av skadenivået på talesystemet, er de delt inn i:
1) afasi - forfall av alle komponenter i tale som følge av skade på kortikale taleområder;
2) alalia - systemisk underutvikling av tale på grunn av lesjoner av kortikale talesoner i pre-taleperioden;
3) dysartri - et brudd på lyduttaleaspektet ved tale som et resultat av et brudd på innerveringen av talemusklene.
Avhengig av plasseringen av lesjonen, skilles flere former for dysartri.
II. Taleforstyrrelser assosiert med funksjonelle endringer
sentralnervesystemet:
1) stamming;
2) mutisme og surdomutisme.
III. Taleforstyrrelser assosiert med defekter i strukturen til artikulasjonsapparatet (mekanisk dyslalia, rhinolalia).
IV. Forsinkelser i taleutvikling av ulik opprinnelse (på grunn av prematuritet, somatisk svakhet, pedagogisk omsorgssvikt, etc.).
Sensorisk afasi(Wernickes afasi), eller verbal "døvhet", oppstår når den venstre temporale regionen er skadet (den midtre og bakre delen av den øvre temporal gyrus). A. R. Luria skiller to former for sensorisk afasi: akustisk-gnostisk og akustisk-mnestisk.
Grunnlaget for defekten akustisk-gnostisk form utgjør et brudd på auditiv gnosis. Pasienten skiller seg ikke ved å høre fonemer som er like i lyd i fravær av døvhet (fonemisk analyse vurderes), som et resultat av at forståelsen av betydningen av individuelle ord og setninger blir forvrengt og svekket. Alvorlighetsgraden av disse lidelsene kan variere. I de alvorligste tilfellene blir den adresserte talen ikke oppfattet i det hele tatt og ser ut til å være tale på et fremmedspråk. Denne formen oppstår når den bakre delen av den øvre temporale gyrusen på venstre hjernehalvdel er skadet - Brodmann-område 22.
I hjernebarken analyseres alle stimuli som kommer fra det omkringliggende ytre og indre miljøet. Det største antallet afferente impulser når cellene i 3. og 4. lag av hjernebarken. Cerebral cortex inneholder sentre som regulerer utførelsen av visse funksjoner. I. P. Pavlov betraktet hjernebarken som et sett med kortikale ender av analysatorer. Begrepet "analysator" refererer til et komplekst kompleks av anatomiske strukturer, som består av et perifert reseptor (oppfattende) apparat, ledere av nerveimpulser og et senter. I evolusjonsprosessen er funksjoner lokalisert i hjernebarken. Den kortikale enden av analysatorene er ikke noen strengt definert sone. I hjernebarken skilles en "kjerne" av sansesystemet og "spredte elementer". Kjernen er området hvor det største antallet kortikale nevroner, der alle strukturer av den perifere reseptoren er nøyaktig projisert. Spredte elementer er plassert nær kjernen og videre ulike avstander Fra han. Hvis høyere analyse og syntese utføres i kjernen, utføres enklere analyser i spredte elementer. Samtidig har ikke sonene med "spredte elementer" til forskjellige analysatorer klare grenser og overlapper hverandre.
Funksjonelle egenskaper av de kortikale sonene i frontallappen. I området til den presentrale gyrusen til frontallappen er det den kortikale kjernen til motoranalysatoren. Dette området kalles også sensorimotorisk cortex. Noen av de afferente fibrene fra thalamus kommer hit og bærer proprioseptiv informasjon fra kroppens muskler og ledd (fig. 8.7). Nedadgående veier til hjernestammen og ryggmargen begynner også her, noe som gir mulighet for bevisst regulering av bevegelser (pyramideformede kanaler). Skader på dette området av cortex fører til lammelse av den motsatte halvdelen av kroppen.
Ris. 8.7. Somatotopisk fordeling i presentral gyrus
Sentrum av skrift ligger i den bakre tredjedelen av den midtre frontale gyrusen. Denne sonen av cortex gir projeksjoner til kjernene til de okulomotoriske kraniale nervene, og kommuniserer også, gjennom kortiko-kortikale forbindelser, med synssenteret i occipitallappen og kontrollsenteret for musklene i armer og nakke i presentralen. gyrus. Skader på dette senteret fører til svekkede skriveferdigheter under visuell kontroll (agrafi).
Talemotorsenteret (Brocas sentrum) ligger i området til den nedre frontale gyrusen. Den har uttalt funksjonell asymmetri. Når den blir ødelagt i høyre hjernehalvdel, går evnen til å regulere klang og intonasjon tapt, talen blir monoton. Når det talemotoriske senteret til venstre blir ødelagt, blir taleartikulasjonen irreversibelt svekket, opp til tap av evnen til å artikulere tale (afasi) og sang (amusia). Med delvise brudd kan agrammatisme observeres - manglende evne til å danne setninger riktig.
I regionen av fremre og midtre tredjedel av øvre, midtre og delvis nedre frontale gyri er det en omfattende fremre assosiativ sone av cortex, som programmerer komplekse former for atferd (planlegging ulike former aktiviteter, beslutningstaking, analyse av oppnådde resultater, frivillig forsterkning av aktiviteter, korrigering av motivasjonshierarkiet).
Området til frontalpolen og medial frontal gyrus er assosiert med reguleringen av aktiviteten til emosjoniske områder av hjernen inkludert i det limbiske systemet, og er relatert til kontrollen av psyko-emosjonelle tilstander. Forstyrrelser i dette området av hjernen kan føre til endringer i det som vanligvis kalles "personlighetsstruktur" og påvirke en persons karakter, hans verdiorientering og intellektuell aktivitet.
Orbitalregionen inneholder sentrene til luktanalysatoren og er nært forbundet anatomisk og funksjonelt med det limbiske systemet i hjernen.
Funksjonelle egenskaper av de kortikale sonene i parietallappen. I den postsentrale gyrusen og den øvre parietale lobulen er det det kortikale senteret til analysatoren for generell sensitivitet (smerte, temperatur og taktil), eller somatosensorisk cortex. Representasjonen av forskjellige deler av kroppen i den, som i den presentrale gyrusen, er bygget i henhold til det somatotopiske prinsippet. Dette prinsippet forutsetter at kroppsdeler projiseres på overflaten av sporet i de topografiske forhold som de har i menneskekroppen. Representasjonen av ulike deler av kroppen i hjernebarken varierer imidlertid betydelig. Den største representasjonen er de områdene (hånd, hode, spesielt tunge og lepper) som er assosiert med komplekse bevegelser som skrift, tale osv. Krenkelser av cortex i dette området fører til delvis eller fullstendig anestesi (tap av følsomhet).
Lesjoner av cortex i området av den overordnede parietallobuen fører til en reduksjon i smertefølsomhet og svekkelse av stereognose - gjenkjenning av gjenstander ved berøring uten hjelp av syn.
I den nedre parietale lobulen, i regionen til den supramarginale gyrusen, er det et senter for praxia, som regulerer evnen til å utføre komplekst koordinerte handlinger som danner grunnlaget for arbeidsprosesser, som krever spesiell opplæring. Et betydelig antall nedadgående fibre som følger som en del av banene som styrer bevisste bevegelser (pyramideformede baner) stammer også herfra. Dette området av parietal cortex, gjennom kortiko-kortikale forbindelser, samhandler tett med frontal cortex og med alle sensoriske områder i den bakre hjernehalvdelen.
Det visuelle (optiske) talesenteret er lokalisert i vinkelgyrusen til parietallappen. Dens skade fører til manglende evne til å forstå lesbar tekst (alexia).
Funksjonelle egenskaper ved de kortikale sonene i occipitallappen. I området av calcarine sulcus er det kortikale sentrum av den visuelle analysatoren. Dens skade fører til blindhet. Hvis det er forstyrrelser i områdene av cortex ved siden av calcarine sulcus i området av den oksipitale polen på de mediale og laterale overflatene av lappen, kan tap av visuell hukommelse, evnen til å navigere i et ukjent miljø oppstå, funksjoner forbundet med med binokulært syn forstyrres (evnen til å bruke synet til å evaluere formen på objekter, avstanden til dem, korrekt proporsjonere bevegelser i rommet under visuell kontroll, etc.).
Funksjonelle egenskaper av de kortikale sonene i tinninglappen. I området til den overordnede temporale gyrusen, dypt i lateral sulcus, er det kortikale sentrum av den auditive analysatoren. Dens skade fører til døvhet.
Det auditive talesenteret (Wernickes sentrum) ligger i den bakre tredjedelen av den øvre temporal gyrus. Skader i dette området resulterer i manglende evne til å forstå muntlig tale: det oppfattes som støy (sensorisk afasi).
I området av den midtre og nedre temporale gyri er det en kortikal representasjon av den vestibulære analysatoren. Skader på dette området fører til ubalanse ved stående og nedsatt følsomhet av det vestibulære apparatet.
Funksjonelle egenskaper av de kortikale sonene i insulaen.
Informasjon om funksjonene til insulaen er motstridende og utilstrekkelig. Det er bevis på at cortex i den fremre delen av insulaen er relatert til analysen av lukt- og smaksopplevelser, og den bakre delen er relatert til prosessering av somatosensorisk informasjon og auditiv oppfatning av tale.
Funksjonelle egenskaper ved det limbiske systemet. Det limbiske systemet– et sett av en rekke hjernestrukturer, inkludert cingulate gyrus, isthmus, dentate og parahippocampal gyri, etc. Deltar i reguleringen av funksjonene til indre organer, lukt, instinktiv atferd, følelser, hukommelse, søvn, våkenhet, etc.
Den cingulate og parahippocampale gyri er direkte relatert til det limbiske systemet i hjernen (fig. 8.8 og 8.9). Den kontrollerer et kompleks av vegetative og atferdsmessige psyko-emosjonelle reaksjoner på ytre miljøpåvirkninger. Den kortikale representasjonen av smaks- og luktanalysatorene er lokalisert i parahippocampus gyrus og uncus. Samtidig spiller hippocampus en viktig rolle i læring: mekanismene for korttids- og langtidshukommelse er forbundet med den.

Ris. 8.8. Medial overflate av hjernen
Basal (subkortikale sentrale) kjerner – ansamlinger av grå substans som danner separat liggende kjerner som ligger nærmere bunnen av hjernen. Disse inkluderer striatum, som utgjør den dominerende massen av halvkulene hos lavere virveldyr; gjerde og amygdala (Fig. 8.10).

Ris. 8.9. Det limbiske systemet

Ris. 8.10. Basalganglier
Striatum består av caudate og lentikulære kjerner. Den grå substansen til caudate og lentikulære kjerner veksler med lag av hvit substans, noe som førte til det vanlige navnet på denne gruppen av subkortikale kjerner - striatum.
Caudatkjernen er lokalisert lateralt og overlegen thalamus, og er atskilt fra den av stria terminalis. Caudatkjernen har hode, kropp og hale. Lentikulærkjernen er lokalisert lateralt for caudatum. Et lag med hvit substans, den indre kapselen, skiller linsekjernen fra kaudatet og fra thalamus. I den lentikulære kjernen skilles globus pallidus (medialt) og putamen (lateralt). Den ytre kapselen (en smal stripe av hvit substans) skiller skallet fra kabinettet.
Caudatikjernen, putamen og globus pallidus kontrollerer komplekst koordinerte automatiserte bevegelser av kroppen, kontrollerer og opprettholder tonen i skjelettmuskulaturen, og er også det høyeste senteret for regulering av slike autonome funksjoner som varmeproduksjon og karbohydratmetabolisme i musklene i kropp. Hvis putamen og globus pallidus er skadet, kan langsomme, stereotype bevegelser (athetose) observeres.
Kjernene i striatum tilhører det ekstrapyramidale systemet, som er involvert i kontroll av bevegelser og regulering av muskeltonus.
Gjerdet er en vertikal plate av grå substans, hvis nedre del fortsetter inn i substansen til den fremre perforerte platen ved bunnen av hjernen. Gjerdet er plassert i den hvite substansen i halvkulen lateralt for linsekjernen og har mange forbindelser med hjernebarken.
Amygdala ligger i den hvite substansen i tinninglappen på halvkulen, 1,5–2 cm bak tinningpolen, gjennom kjernene har den forbindelser med hjernebarken, med strukturene i luktesystemet, med hypothalamus og kjernene. av hjernestammen som kontrollerer kroppens autonome funksjoner. Dens ødeleggelse fører til aggressiv oppførsel eller en apatisk, sløv tilstand. Gjennom sine forbindelser med hypothalamus påvirker amygdala det endokrine systemet så vel som reproduktiv atferd.
Den hvite substansen i halvkulen inkluderer den indre kapselen og fibrene som passerer gjennom hjernekommissurene (corpus callosum, anterior commissur, fornix commissur) og går til cortex og basalganglia, fornix, samt systemer av fibre som forbinder områder av cortex og subkortikale sentre innenfor den ene halvdelen av hjernen (hemisfærer).
I og II laterale ventrikler. Hulrommene i hjernehalvdelene er sideventriklene (I og II), som ligger i tykkelsen av den hvite substansen under corpus callosum. Hver ventrikkel består av fire deler: det fremre hornet ligger i frontalen, den sentrale delen - i parietalen, det bakre hornet - i det occipitale og det nedre hornet - i tinninglappen (fig. 8.11).
De fremre hornene til begge ventriklene er atskilt fra hverandre av to plater med en gjennomsiktig skillevegg. Den sentrale delen av sideventrikkelen bøyer seg ovenfra rundt thalamus, danner en bue og passerer bakover - inn i det bakre hornet, nedover i det nedre hornet. Choroid plexus stikker ut i den sentrale delen og nedre horn av lateral ventrikkel, som kobles til choroid plexus i tredje ventrikkel gjennom interventrikulære foramen.

Ris. 8.11. Hjernens ventrikler:
1 - venstre hjernehalvdel, 2 - laterale ventrikler, 3 - tredje ventrikkel, 4 - midthjerneakvedukt, 5 - fjerde ventrikkel, 6 - lillehjernen, 7 - inngang til den sentrale kanalen i ryggmargen, 8 - ryggmarg
Ventrikkelsystemet inkluderer parvise C-formede hulrom - de laterale ventriklene med deres fremre, nedre og bakre horn, som strekker seg henholdsvis inn i frontallappene, tinninglappene og occipitallappene i hjernehalvdelene. Omtrent 70 % av all cerebrospinalvæske skilles ut av choroid plexus i veggene i sideventriklene.
Fra de laterale ventriklene passerer væske gjennom den interventrikulære foramina inn i det spaltelignende hulrommet i den tredje ventrikkelen, som ligger i hjernens sagittale plan og deler thalamus og hypothalamus i to symmetriske halvdeler. Hulrommet til den tredje ventrikkelen er forbundet med en smal kanal - akvedukten til midthjernen (akvedukten til Sylvius) med hulrommet til den fjerde ventrikkelen. Den fjerde ventrikkelen kommuniserer gjennom flere kanaler (åpninger) med subaraknoidalrommene i hjernen og ryggmargen.
Diencephalon
Diencephalon ligger under corpus callosum og består av thalamus, epithalamus, metathalamus og hypothalamus (fig. 8.12, se fig. 7.2).
Thalamus(visuell tuberkel) – sammenkoblet, eggformet, hovedsakelig dannet av grå substans. Thalamus er det subkortikale senteret for alle typer følsomhet. Den mediale overflaten av høyre og venstre thalami, vendt mot hverandre, danner sideveggene i hulrommet til diencephalon - den tredje ventrikkelen; de er forbundet med hverandre ved en interthalamusfusjon. Thalamus inneholder grå substans, som består av klynger av nevroner som danner thalamuskjernene. Kjernene er atskilt av tynne lag med hvitt stoff. Omtrent 40 kjerner av thalamus ble studert. Hovedkjernene er fremre, mediale, bakre.

Ris. 8.12. Hjernedeler
Epithalamus inkluderer pinealkjertelen, bånd og båndtrekanter. Pinealkroppen, eller pinealkjertelen, som er en endokrin kjertel, er så å si opphengt i to bånd, forbundet med en kommissur og koblet til thalamus gjennom trekanter av bånd. Båndtrekantene inneholder kjerner relatert til lukteanalysatoren. Hos en voksen er den gjennomsnittlige lengden på epifysen ~0,64 cm og massen er ~0,1 g. Metathalamus dannet av parede mediale og laterale genikulære kropper som ligger bak hver thalamus. Den mediale genikulære kroppen er plassert bak thalamusputen; den er, sammen med de nedre colliculi av midthjernens takplate (quadrigeminal), det subkortikale sentrum av den auditive analysatoren. Lateral - plassert nedover fra puten, er den, sammen med de øvre colliculi på takplaten, det subkortikale sentrum av den visuelle analysatoren. Kjerner genikulerte kropper knyttet til de kortikale sentrene til de visuelle og auditive analysatorene.
Hypothalamus, som representerer den ventrale delen av diencephalon, er plassert foran de cerebrale peduncles og inkluderer en rekke strukturer som har ulike opphav– den fremre lokaliserte visuelle delen er dannet fra telencephalon (optisk chiasme, optisk trakt, grå tuberkel, infundibulum, neurohypophysis); fra den mellomliggende - den olfaktoriske delen (mammillære kropper og selve subthalamus-regionen - hypothalamus) (fig. 8.13).

Figur 8.13. Basalganglier og diencephalon
Hypothalamus er senteret for regulering av endokrine funksjoner; den kombinerer nervøse og endokrine reguleringsmekanismer til et felles nevroendokrine system, koordinerer nervøse og hormonelle mekanismer for å regulere funksjonene til indre organer. Hypothalamus inneholder nevroner av vanlig type og nevrosekretoriske celler. Hypothalamus og hypofysen danner et enkelt funksjonelt kompleks, der førstnevnte spiller en regulerende og sistnevnte en effektorrolle.
Hypothalamus har mer enn 30 par kjerner. Store nevrosekretoriske celler i de supraoptiske og paraventrikulære kjernene i den fremre hypotalamiske regionen produserer nevrosekreter av peptidnatur.
Den mediale hypothalamus inneholder nevroner som oppfatter alle endringer som skjer i blodet og cerebrospinalvæsken (temperatur, sammensetning, hormoninnhold, etc.). Den mediale hypothalamus er også koblet til den laterale hypothalamus. Sistnevnte har ikke kjerner, men har bilaterale forbindelser med de overliggende og underliggende delene av hjernen. Den mediale hypothalamus er en kobling mellom nervesystemet og det endokrine systemet. De siste årene har enkefaliner og endorfiner (peptider), som har en morfinlignende effekt, blitt isolert fra hypothalamus. De antas å være involvert i regulering av atferd og vegetative prosesser.
Foran den bakre perforerte substansen ligger to små sfæriske mastoidlegemer, dannet av grå substans dekket med et tynt lag av hvitt. Kjernene til mammillærlegemene er de subkortikale sentrene til lukteanalysatoren. Foran mastoidlegemene er en grå tuberkel, som er begrenset foran av den optiske chiasmen og den optiske kanalen; det er en tynn plate av grå substans i bunnen av den tredje ventrikkelen, som strekker seg nedover og fremad og danner en trakt . Slutten av det går inn hypofysen – en endokrin kjertel som ligger i hypofysen i sella turcica. Kjernene i det autonome nervesystemet ligger i den grå haugen. De påvirker også en persons følelsesmessige reaksjoner.
Den delen av diencephalon, som ligger under thalamus og atskilt fra den av hypothalamus-sporet, utgjør selve hypothalamus. Dekkene til hjernestammene fortsetter her, de røde kjernene og den svarte substansen i mellomhjernen slutter her.
III ventrikkel. Diencephalons hulrom - III ventrikkel Det er et smalt spaltelignende rom som ligger i sagittalplanet, avgrenset lateralt av de mediale overflatene av thalamus, under av hypothalamus, foran av søylene i fornix, den fremre kommissur og terminalplaten, bak av epithalamus (bakre) kommissur, over av fornix, over hvilken corpus callosum er plassert. Den øvre veggen i seg selv er dannet av den vaskulære basen til den tredje ventrikkelen, der dens choroid plexus ligger.
Kaviteten til den tredje ventrikkelen går bakover inn i midthjerneakvedukten, og foran på sidene kommuniserer den interventrikulære foramina med laterale ventrikler.
Midthjerne
Mellomhjernen – den minste delen av hjernen, som ligger mellom diencephalon og pons (fig. 8.14 og 8.15). Området over akvedukten kalles taket av midthjernen, og på det er det fire konveksiteter - den quadrigeminale platen med de overlegne og underordnede colliculi. Det er her de visuelle og auditive refleksbanene går til ryggmargen.
Cerebrale peduncles er hvite runde snorer som kommer ut av pons og beveger seg fremover til hjernehalvdelene. Den oculomotoriske nerven (III par kraniale nerver) kommer ut av sporet på den mediale overflaten av hver peduncle. Hvert ben består av et dekk og en base, grensen mellom dem er en svart substans. Fargen avhenger av overflod av melanin i nervecellene. Den substantia nigra tilhører det ekstrapyramidale systemet, som er involvert i å opprettholde muskeltonus og automatisk regulerer muskelfunksjonen. Basen av pedikelen er dannet av nervetråder som går fra hjernebarken til ryggmargen og medulla oblongata og pons. Tegmentum av hjernestammene inneholder hovedsakelig stigende fibre på vei til thalamus, blant hvilke kjernene ligger. De største er de røde kjernene, hvorfra den motoriske røde kjerne-spinalkanalen begynner. I tillegg er den retikulære formasjonen og kjernen til den dorsale longitudinelle fasciculus (mellomkjerne) lokalisert i tegmentum.
bakhjerne
Bakhjernen inkluderer de ventralt plasserte pons og lillehjernen som ligger bak pons.

Ris. 8.14. Skjematisk fremstilling av et lengdesnitt av hjernen

Ris. 8.15. Tverrsnitt gjennom midthjernen på nivå med colliculus superior (planet til snittet er vist i fig. 8.14)
Bro ser ut som en liggende tverrgående fortykket ås, fra sidesiden av hvilken de midtre lillehjernens peduncles strekker seg til høyre og venstre. Den bakre overflaten av pons, dekket av lillehjernen, deltar i dannelsen av rhomboid fossa, den fremre overflaten (tilstøtende bunnen av hodeskallen) grenser til medulla oblongata nedenfor og cerebrale peduncles ovenfor (se fig. 8.15). Den er tverrstripet på grunn av den tverrgående retningen til fibrene som går fra pontinkjernene til de midtre lillehjernens peduncles. På den fremre overflaten av broen langs midtlinjen er det et basilarspor plassert i lengderetningen, der arterien med samme navn passerer.
Broen består av mange nervefibre som danner veier, blant annet cellulære klynger - kjerner. De fremre banene forbinder cerebral cortex med ryggmargen og cerebellar cortex. I den bakre delen av broen (tegmentum) er det stigende veier og delvis synkende, den retikulære formasjonen, kjernene til V, VI, VII, VIII parene av kranienerver er lokalisert. På grensen mellom begge deler av broen ligger et trapesformet legeme dannet av kjernene og tverrgående fibre i den ledende banen til den auditive analysatoren.
Lillehjernen spiller en stor rolle i å opprettholde kroppens balanse og koordinering av bevegelser. Lillehjernen når sin største utvikling hos mennesker i forbindelse med oppreist holdning og tilpasning av hånden til arbeid. I denne forbindelse har mennesker høyt utviklede halvkuler (ny del) av lillehjernen.
I lillehjernen er det to halvkuler og en uparret midtre fylogenetisk gammel del - vermis (fig. 8.16).

Ris. 8.16. Lillehjernen: topp- og bunnvisning
Overflatene på halvkulene og vermis er atskilt av tverrgående parallelle riller, mellom hvilke det er smale lange blader av lillehjernen. Lillehjernen er delt inn i fremre, bakre og floculonodulære lapper, atskilt av dypere sprekker.
Lillehjernen består av grå og hvit substans. Den hvite substansen, som trenger inn mellom den grå substansen, ser ut til å forgrene seg, og danner på den midtre delen figuren av et forgrenet tre - "livets tre" i lillehjernen.
Cerebellar cortex består av grå substans 1–2,5 mm tykk. I tillegg, i tykkelsen av den hvite substansen er det ansamlinger av grå - parede kjerner: dentate kjerne, korkformet, sfærisk og telt kjerne. Afferente og efferente fibre som forbinder cerebellum med andre deler danner tre par cerebellare peduncles: de nedre går til medulla oblongata, de midterste til pons, de øvre til quadrigemulus.
Ved fødselen er lillehjernen mindre utviklet enn telencephalon (spesielt halvkulen), men i det første leveåret utvikler den seg raskere enn andre deler av hjernen. En uttalt utvidelse av lillehjernen observeres mellom 5. og 11. levemåned, når barnet lærer å sitte og gå.
Medulla er en direkte fortsettelse av ryggmargen. Dens nedre grense anses å være utgangsstedet for røttene til den første cervikale spinalnerven eller dekussjonen av pyramidene, den øvre er den bakre kanten av broen, lengden er omtrent 25 mm, formen nærmer seg en avkortet kjegle , med basen vendt oppover.
Den fremre overflaten er delt av den fremre medianfissuren, på sidene av hvilke det er pyramider dannet av pyramideformede baner som delvis krysser hverandre (pyramidedekussering) i dybden av den beskrevne sprekken ved grensen til ryggmargen. Fibre i de pyramidale kanalene forbinder hjernebarken med kjernene til kranienervene og de fremre hornene i ryggmargen. På hver side av pyramiden er det en oliven, adskilt fra pyramiden av den fremre sidesporet.
Den bakre overflaten av medulla oblongata er delt av den bakre median sulcus; på hver side av den er det fortsettelser av de bakre ledningene av ryggmargen, som divergerer oppover og går inn i de underordnede cerebellare peduncles.
Medulla oblongata er bygget av hvitt og grått stoff, sistnevnte er representert av kjernene til IX–XII parene av kranienerver, oliven, sentre for respirasjon og sirkulasjon, og retikulær formasjon. Hvit substans er dannet av lange og korte fibre som utgjør de tilsvarende banene.
Retikulær formasjon er en samling av celler, celleklynger og nervefibre som ligger i hjernestammen (medulla oblongata, pons og midthjernen) og danner et nettverk. Den retikulære formasjonen er koblet til alle sanseorganer, motoriske og sensoriske områder i hjernebarken, thalamus og hypothalamus, og ryggmargen. Den regulerer nivået av eksitabilitet og tonus i ulike deler av sentralnervesystemet, inkludert hjernebarken, og er involvert i reguleringen av bevissthetsnivået, følelser, søvn og våkenhet, autonome funksjoner og målrettede bevegelser.
IV ventrikkel- Dette er hulrommet i rombehjernen; nedover fortsetter det inn i den sentrale kanalen i ryggmargen. Bunnen av IV-ventrikkelen kalles på grunn av formen en romboid fossa (fig. 8.17). Den er dannet av de bakre overflatene av medulla oblongata og pons, de øvre sidene av fossaen er de øvre, og de nedre er de underordnede cerebellar peduncles.

Ris. 8.17. Hjernestamme; bakfra. Lillehjernen fjernes, rhomboid fossa er åpen
Mediansporet deler bunnen av fossaen i to symmetriske halvdeler; på begge sider av sporet er mediale forhøyninger synlige, som utvider seg midt i fossaen inn i høyre og venstre ansikts tuberkler, hvor de ligger: kjernen til VI par kranialnerver (abducens nerve), dypere og mer lateral - kjernen til VII-paret (ansiktsnerven), og nedover går den mediale eminensen inn i trekanten til hypoglossal nerve, lateralt som er trekanten til vagusnerven. I trekantene, i tykkelsen av hjernestoffet, ligger kjernene til nervene med samme navn. Den overordnede vinkelen til rhomboid fossa kommuniserer med midthjerneakvedukten. De laterale seksjonene av rhomboid fossa kalles vestibulære felt, hvor de auditive og vestibulære kjernene til vestibulocochlear nerve (VIII par kraniale nerver) ligger. Fra de auditive kjernene strekker seg tverrgående margstriper til median sulcus, som ligger på grensen mellom medulla oblongata og pons og er fibrene i den ledende banen til auditiv analysator. I tykkelsen av rhomboid fossa ligger kjernene til V, VI, VII, VIII, IX, X, XI og XII parene av kranienerver.
Blodtilførsel til hjernen
Blod kommer inn i hjernen gjennom to parede arterier: den indre carotis og vertebral. I kraniehulen smelter begge vertebrale arterier sammen, og danner sammen hovedarterien (basal). Ved bunnen av hjernen går basilarterien sammen med de to halspulsårene, og danner en enkelt arteriell ring (fig. 8.18). Denne kaskademekanismen for blodtilførsel til hjernen sikrer tilstrekkelig blodstrøm hvis noen av arteriene svikter.

Ris. 8.19. Arterier ved bunnen av hjernen og sirkelen til Willis ( høyre hjernehalvdel lillehjernen og høyre tinninglapp fjernet); Sirkelen til Willis er vist med en stiplet linje
Tre kar går fra den arterielle ringen: de fremre, bakre og midtre cerebrale arteriene, som forsyner hjernehalvdelene. Disse arteriene går langs overflaten av hjernen, og fra dem blir blod levert dypt inn i hjernen av mindre arterier.
Halspulsåresystemet kalles carotissystemet, som gir 2/3 av hjernens arterielle blodbehov og forsyner de fremre og midtre delene av hjernen.
Det "vertebrale-basale" arteriesystemet kalles det vertebrobasilære systemet, som gir 1/3 av hjernens behov og leverer blod til de bakre delene.
Utstrømningen av veneblod skjer hovedsakelig gjennom de overfladiske og dype cerebrale venene og venøse bihulene (fig. 8.19). Blodet strømmer til slutt inn i den indre halsvenen, som kommer ut av skallen gjennom halshulen, som ligger ved bunnen av skallen lateralt for foramen magnum.
Meninges
Hjernens membraner beskytter den mot mekanisk skade og fra penetrasjon av infeksjoner og giftige stoffer (fig. 8.20).

Ris. 8.19. Vener og venøse bihuler i hjernen

Fig.8.20. Koronal snitt gjennom skalleskallet og hjernen
Den første membranen som beskytter hjernen kalles pia mater. Det er tett ved siden av hjernen, strekker seg inn i alle rillene og hulrommene (ventriklene) som er tilstede i tykkelsen av selve hjernen. Hjernens ventrikler er fylt med en væske som kalles cerebrospinalvæske eller cerebrospinalvæske. Dura mater er direkte ved siden av beinene i skallen. Mellom de myke og harde membranene er arachnoid (arachnoid) membranen. Mellom arachnoid og myke membraner er det et rom (subaraknoidal eller subaraknoidalrom) fylt med cerebrospinalvæske. Den arachnoidale membranen sprer seg over rillene i hjernen og danner en bro, og den myke smelter sammen med dem. På grunn av dette dannes hulrom kalt sisterne mellom de to skjellene. Cisternene inneholder cerebrospinalvæske. Disse tankene beskytter hjernen mot mekaniske skader, og fungerer som "kollisjonsputer".
Nerveceller og blodkar er omgitt av neuroglia - spesielle cellulære formasjoner som utfører beskyttende, støttende og metabolske funksjoner, gir de reaktive egenskapene til nervevev og deltar i dannelsen av arr, inflammatoriske reaksjoner, etc.
Når hjernen er skadet, aktiveres plastisitetsmekanismen, når de gjenværende hjernestrukturene overtar funksjonene til de berørte områdene.
Deretter var innsatsen til fysiologer rettet mot å søke etter "kritiske" områder av hjernen, hvis ødeleggelse førte til forstyrrelse av refleksaktiviteten til et bestemt organ. Gradvis dukket ideen om en stiv anatomisk lokalisering av "refleksbuer" opp, og følgelig begynte refleksen i seg selv å bli tenkt på som en mekanisme for drift av bare de nedre delene av hjernen (spinalsentre).
Samtidig ble spørsmålet om lokalisering av funksjoner i de høyere delene av hjernen utviklet. Ideer om lokalisering av elementer av mental aktivitet i hjernen oppsto for lenge siden. I nesten hver epoke, visse eller
Andre hypoteser for representasjon av høyere mentale funksjoner og bevissthet i hjernen generelt.
Østerriksk lege og anatom Franz Joseph Gall(1758-1828) utgjorde Detaljert beskrivelse anatomi og fysiologi av det menneskelige nervesystemet, utstyrt med et utmerket atlas.
: En hel generasjon forskere bygget på disse dataene. Blant Galls anatomiske funn er følgende: identifikasjon av hovedforskjellene mellom den grå og hvite substansen i hjernen; bestemmelse av opprinnelsen til nerver i grå substans; definitive bevis på omtalen av pyramidale kanaler og optiske nerver; etablering av forskjeller mellom "konvergent" (i moderne terminologi "assosiativ") og "divergent" ("projeksjon") fibre (1808); første klare beskrivelse av hjernekommissurer; bevis på begynnelsen av kranienervene i medulla oblongata (1808), etc. Gall var en av de første som tildelte hjernebarken en avgjørende rolle i hjernens funksjonelle aktivitet. Dermed mente han at foldingen av hjerneoverflaten er en utmerket løsning av natur og evolusjon på et viktig problem: å sikre en maksimal økning i hjernens overflate, samtidig som volumet opprettholdes mer eller mindre konstant. Gall introduserte begrepet "bue", kjent for enhver fysiolog, og beskrev dens klare inndeling i tre deler.
Galls navn er imidlertid hovedsakelig kjent i forbindelse med hans ganske tvilsomme (og noen ganger skandaløse!) doktrine om lokalisering av høyere mentale funksjoner i hjernen. Ved å legge stor vekt på samsvaret mellom funksjon og struktur, ga Gall tilbake i 1790 en forespørsel om å introdusere en ny vitenskap i arsenalet av kunnskap - frenologi(fra den greske phren - sjel, sinn, hjerte), som også fikk et annet navn - psykomorfologi, eller smal lokalisering. Som lege observerte Gall pasienter med ulike forstyrrelser i hjerneaktiviteten og la merke til at sykdommens spesifikasjoner i stor grad var avhengig av hvilken del av hjernen som var skadet. Dette førte ham til ideen om at hver mental funksjon tilsvarer en spesiell del av hjernen. Etter å ha sett den endeløse variasjonen av karakterer og individuelle mentale kvaliteter til mennesker, foreslo Gall at styrkingen (eller større overvekt) i menneskelig oppførsel av enhver karaktertrekk eller mental funksjon innebærer foretrukket utvikling av et bestemt område av hjernebarken hvor denne funksjonen er representert. Dermed ble oppgaven fremmet: funksjon lager struktur. Som et resultat av veksten av dette hypertrofierte området av cortex ("hjernekjeglen"), øker trykket på beinene i skallen, noe som igjen forårsaker utseendet til en ekstern kranial tuberkel over det tilsvarende området av hjernen. Ved underutvikling av funksjonen, omvendt.
En merkbar depresjon ("grop") vil vises på overflaten av hodeskallen. Ved å bruke metoden for "kranioskopi" laget av Gall - å studere lettelsen av hodeskallen ved hjelp av palpasjon - og detaljerte "topografiske" kart over overflaten av hjernen, som indikerte plasseringen av alle evner (betraktet som medfødt), laget Gall og hans tilhengere en diagnose, dvs. gjort en konklusjon om karakter og tilbøyeligheter til en person, om hans mentale og moralske egenskaper. Ble det tildelt 2? områder av hjernen hvor visse evner til et individ er lokalisert (og 19 av dem ble anerkjent som felles for mennesker og dyr, og 8 som rent menneskelige). I tillegg til "humpene" som er ansvarlige for implementeringen av fysiologiske funksjoner, var det også de som vitnet om visuell og auditiv hukommelse, orientering i rommet, en følelse av tid og instinktet for forplantning; slik personlige kvaliteter. slik som mot, ambisjoner, fromhet, vidd, hemmelighold, amorøsitet, forsiktighet, selvtillit, raffinement, håp, nysgjerrighet, tilgjengelighet for utdanning, stolthet, uavhengighet, flid, aggressivitet, troskap, kjærlighet til livet, kjærlighet til dyr.
Galls feilaktige og pseudovitenskapelige ideer (som imidlertid var ekstremt populære på hans tid) inneholdt et rasjonelt korn: erkjennelse av den nære sammenhengen mellom manifestasjonene av mentale funksjoner og aktiviteten til hjernebarken. Problemet med å finne differensierte «hjernesentre» og rette oppmerksomheten mot hjernens funksjoner ble satt på dagsorden. Gall kan virkelig betraktes som grunnleggeren av "cerebral lokalisering." Selvfølgelig, for den videre utviklingen av psykofysiologi, var det mer lovende å stille et slikt problem enn det eldgamle søket etter plasseringen av den "vanlige sensoriske".
Løsningen på spørsmålet om lokalisering av funksjoner i hjernebarken ble tilrettelagt av data akkumulert i klinisk praksis og i dyreforsøk. Tysk lege, anatom og fysiker Julius Robert Mayer(1814-1878), som i lang tid praktiserte i parisiske klinikker, og også tjente som skipslege, observerte hos pasienter med traumatiske hjerneskader avhengigheten av svekkelse (eller fullstendig tap) av en eller annen funksjon på skade på en bestemt område av hjernen. Dette tillot ham å foreslå at hukommelsen er lokalisert i hjernebarken (det bør bemerkes at T. Willis kom til en lignende konklusjon tilbake på 1600-tallet), fantasi og dømmekraft er lokalisert i den hvite substansen i hjernen, apperepsjon og vilje er lokalisert i basalgangliene. I følge Mayer er et slags "integrert organ" for atferd og psyke corpus callosum og lillehjernen.
Over tid ble kliniske studier av konsekvensene av hjerneskade supplert med laboratorier kunstig ekstirpasjonsmetode(fra latin ex(s)tirpatio - fjerning med rot), som gjør det mulig å delvis eller fullstendig ødelegge (fjerne) områder av hjernen til dyr for å bestemme deres funksjonelle rolle i hjerneaktivitet. På begynnelsen av 1800-tallet. De utførte hovedsakelig akutte eksperimenter på dyr (frosker, fugler); senere, med utviklingen av asepsismetoder, begynte de å utføre kroniske eksperimenter, som gjorde det mulig å observere dyrs oppførsel i mer eller mindre lang tid etter operasjonen . Fjerning av ulike deler av hjernen (inkludert hjernebarken) hos pattedyr (katter, hunder, aper) gjorde det mulig å belyse det strukturelle og funksjonelle grunnlaget for komplekse atferdsreaksjoner.
Det viste seg at å frata dyr de høyere delene av hjernen (fugler - forhjernen, pattedyr - hjernebarken) generelt ikke forårsaket forstyrrelser av de grunnleggende funksjonene: respirasjon, fordøyelse, utskillelse, blodsirkulasjon, metabolisme og energi. Dyrene beholdt evnen til å bevege seg og reagere på visse ytre påvirkninger. Følgelig skjer reguleringen av disse fysiologiske manifestasjonene av vital aktivitet på lavere nivåer (sammenlignet med hjernebarken) i hjernen. Men når de høyere delene av hjernen ble fjernet, skjedde det dyptgripende endringer i oppførselen til dyr: de ble praktisk talt blinde og døve, "dumme"; de mistet tidligere ervervede ferdigheter og kunne ikke utvikle nye, kunne ikke tilstrekkelig navigere i miljøet, skilte ikke og kunne ikke skille objekter i det omkringliggende rommet. Kort sagt, dyr ble "levende automater" med monotone og ganske primitive måter å reagere på.
I forsøk med delvis fjerning av områder av hjernebarken ble det oppdaget at hjernen er funksjonelt heterogen og ødeleggelse av et eller annet område fører til forstyrrelse av en viss fysiologisk funksjon. Dermed viste det seg at de oksipitale områdene av cortex er assosiert med visuell funksjon, de temporale områdene med auditiv funksjon, området til sigmoid gyrus med motorisk funksjon, samt med hud- og muskelfølsomhet. Dessuten forbedres denne differensieringen av funksjoner i individuelle områder av de høyere delene av hjernen evolusjonær utvikling dyr.
Strategien for vitenskapelig forskning i studiet av hjernefunksjoner førte til at forskere i tillegg til eksstirpasjonsmetoden begynte å bruke metoden for kunstig stimulering av visse områder av hjernen ved hjelp av elektrisk stimulering, noe som også gjorde det mulig å vurdere den funksjonelle rollen til de viktigste delene av hjernen. Dataene oppnådd ved bruk av disse laboratorieforskningsmetodene, samt resultatene av kliniske observasjoner, skisserte en av hovedretningene for psykofysiologi på 1800-tallet. - lokaliseringsbestemmelse nervesentre, ansvarlig for høyere mentale funksjoner og oppførsel av kroppen som helhet. Så. i 1861 uttalte den franske vitenskapsmannen, antropologen og kirurgen Paul Broca (1824-1880), på grunnlag av kliniske fakta, bestemt seg mot den fysiologiske ekvivalensen til hjernebarken. Mens han obduserte likene av pasienter som led av en taleforstyrrelse i form av motorisk afasi (pasientene forsto andres tale, men kunne ikke snakke selv), oppdaget han endringer i den bakre delen av den nedre (tredje) frontale gyrusen til venstre hjernehalvdel eller i den hvite substansen under dette området av cortex. Som et resultat av disse observasjonene etablerte Broca posisjonen til det motoriske (motoriske) talesenteret, senere oppkalt etter ham. I 1874 ble den tyske psykiateren og nevrologen K? Wernicke (1848-1905) beskrev det sensoriske talesenteret (som i dag bærer navnet hans) i den bakre tredjedelen av den første temporale gyrusen på venstre hjernehalvdel. Skader på dette senteret fører til tap av evnen til å forstå menneskelig tale (sensorisk afasi). Enda tidligere, i 1863, ved bruk av metoden for elektrisk stimulering av visse områder av cortex (presentral gyrus, presentral region, fremre del av pericentral lobule, bakre deler av øvre og midterste frontal gyri), tyske forskere Gustav Fritsch og Eduard Hitzig etablerte motoriske sentre (motoriske kortikale felt), hvor irritasjon forårsaket visse sammentrekninger av skjelettmuskulaturen, "og ødeleggelse førte til dype forstyrrelser i motorisk atferd. I 4874 oppdaget Kiev-anatomen og -legen Vladimir Alekseevich Betz (1834-1894) efferente nerveceller av motoriske sentre - gigantiske pyramideceller i cortex lag V, oppkalt etter ham Betz-celler. Den tyske forskeren Hermann Munch (student av J. Müller og E. Dubois-Reymond) oppdaget ikke bare motoriske kortikale felt, ved å bruke eksstirpasjonsmetoden han fant sentre for sanseoppfatninger Han var i stand til å vise at synssenteret er lokalisert i hjernelappen, hørselssenteret er i tinninglappen. Fjerning av oksipitallappen i hjernen førte til tap av dyrets evne til å se (med fullstendig bevaring av det visuelle apparatet). Allerede inne
begynnelsen av det 20. århundre fremragende østerriksk nevrolog Constantin Economo(1876-1931) ble sentrene for svelging og tygging etablert i den såkalte substantia nigra i hjernen (1902), sentrene som kontrollerer søvnen ble funnet i mellomhjernen (1917) Ser vi litt fremover, la oss si at Economo ga en utmerket beskrivelse av strukturen til hjernebarken en voksen og foredlet i 1925 det cytoarkitektoniske kartet over de kortikale feltene i hjernen, og plottet 109 felt på det.
Samtidig skal det bemerkes at på 1800-tallet. Alvorlige argumenter har blitt fremsatt mot posisjonen til smale lokalisasjonister, i henhold til hvis synspunkter motoriske og sensoriske funksjoner er begrenset til forskjellige områder av hjernebarken. Dermed oppsto teorien om ekvivalensen av områder av cortex, og bekreftet ideen om like viktighet av kortikale formasjoner for gjennomføringen av enhver aktivitet i kroppen - ekvipotensialisme. I denne forbindelse ble de frenologiske synspunktene til Gall, en av de mest ivrige tilhengerne av lokalisering, kritisert av den franske fysiologen Marie Jean Pierre Flourens(1794-1867). Tilbake i 1822 påpekte han tilstedeværelsen av et respirasjonssenter i medulla oblongata (som han kalte "vital node"); koblet koordinering av bevegelser med aktiviteten til lillehjernen, syn - med quadrigeminal regionen; Hovedfunksjonen til ryggmargen var å lede eksitasjon langs nervene. Til tross for slike tilsynelatende lokaliseringistiske synspunkter, mente Flourens at de grunnleggende mentale prosessene (inkludert intellekt og vilje) som ligger til grunn for målrettet menneskelig atferd utføres som et resultat av hjernens aktivitet som en integrert formasjon, og derfor kan en integrert atferdsfunksjon ikke assosieres. med en bestemt anatomisk formasjon. Flourens utførte de fleste eksperimentene sine på duer og kyllinger, fjernet individuelle deler av hjernen deres og observerte endringer i fuglenes oppførsel. Fuglenes atferd kom seg vanligvis en stund etter operasjonen, uavhengig av hvilke områder av hjernen som ble skadet, så Flourens konkluderte med at graden av svekkelse av ulike former for atferd først og fremst ble bestemt av hvor mye hjernevev som ble fjernet under operasjonen. Etter å ha forbedret operasjonsteknikken, var han den første som fullstendig fjernet forhjernehalvdelene til dyr og reddet livet deres for videre observasjoner.
Basert på eksperimenter kom Flourens til den konklusjon at forhjernehalvdelene spiller en avgjørende rolle i gjennomføringen av en atferdshandling. Deres fullstendige fjerning fører til tap av alle "intelligente" funksjoner. Dessuten ble spesielt alvorlige atferdsforstyrrelser observert hos kyllinger etter ødeleggelsen av den grå substansen på overflaten av hjernehalvdelene - den såkalte kortikoidplaten, en analog av pattedyrets hjernebark. Flourens foreslo at dette området av hjernen er sete for sjelen, eller "styrende ånd", og derfor fungerer som en enkelt helhet, med en homogen og lik masse (ligner for eksempel på vevsstrukturen i leveren ). Til tross for de litt fantastiske ideene til ekvipotensialistene, er det verdt å merke seg det progressive elementet i deres synspunkter. For det første ble komplekse psykofysiologiske funksjoner gjenkjent som et resultat av den kombinerte aktiviteten til hjerneformasjoner. For det andre ble ideen om høy dynamisk plastisitet i hjernen, uttrykt i utskiftbarheten av delene, fremmet.
- Gall klarte ganske nøyaktig å bestemme "talens sentrum", men det ble "offisielt" oppdaget av den franske forskeren Paul Broca (1861).
- I 1842 kom Mayer, som jobbet med å bestemme den mekaniske ekvivalenten til varme, til en generaliserende lov om bevaring av energi.
- I motsetning til sine forgjengere, som ga nerven evnen til å sanse (dvs. gjenkjenne en viss mental kvalitet bak den), betraktet Hall nerveenden (i sanseorganet) som en "apsykisk" formasjon.
Ideer om lokalisering av funksjoner i hjernebarken er av stor praktisk betydning for å løse problemer med temaet lesjoner i hjernehalvdelene. Imidlertid er mye i denne delen fortsatt kontroversielt og ikke helt løst. Læren om lokalisering av funksjoner i cortex har en ganske lang historie - fra fornektelse av lokalisering av funksjoner i den til fordeling av alle funksjoner i cortex i strengt begrensede territorier menneskelig aktivitet, helt opp til høyeste kvaliteter den siste (minne, vilje, etc.), og til slutt, før du går tilbake til "ekvipotensialiteten" til cortex, det vil si igjen, i hovedsak, til fornektelsen av lokalisering av funksjoner (nylig i utlandet).
Ideer om ekvivalens (ekvipotensialitet) til ulike kortikale felt er i konflikt med det enorme faktamaterialet som er akkumulert av morfologer, fysiologer og klinikere. Daglig klinisk erfaring viser at det er visse urokkelige naturlige avhengigheter av funksjonelle forstyrrelser på plasseringen av det patologiske fokuset. Basert på disse grunnleggende prinsippene løser klinikeren problemene med aktuell diagnostikk. Slik er det imidlertid så lenge vi opererer med lidelser knyttet til relativt enkle funksjoner: bevegelser, følsomhet, etc. Med andre ord er lokalisering i de såkalte "projeksjons"-sonene - kortikale felt, direkte forbundet gjennom deres veier med de underliggende delene av nervesystemet og periferien, godt etablert. Funksjonene til cortex er mer komplekse, fylogenetisk yngre og kan ikke lokaliseres snevert; i gjennomføringen komplekse funksjoner Svært store områder av cortex, og til og med hele cortex som helhet, er involvert. Det er grunnen til at det er vanskeligere og noen ganger unøyaktig å løse problemer med lesjonsemner basert på taleforstyrrelser, apraksi, agnosi og spesielt psykiske lidelser, som klinisk erfaring viser.
Samtidig er det innenfor hjernebarken områder hvis skade forårsaker en eller annen karakter, en eller annen grad, for eksempel taleforstyrrelser, gnoseforstyrrelser og praksi, hvis topodiagnostiske verdi også er betydelig. Det følger imidlertid ikke av dette at det finnes spesielle, snevert lokaliserte sentre som «forvalter» disse mest komplekse formene for menneskelig aktivitet. Det er nødvendig å tydelig skille mellom lokalisering av funksjoner og lokalisering av symptomer.
Grunnlaget for en ny og progressiv doktrine om lokalisering av funksjoner i hjernen ble skapt av I.P. Pavlov.
I stedet for ideen om hjernebarken som til en viss grad en isolert overbygning over andre etasjer i nervesystemet med smalt lokaliserte områder forbundet langs overflaten (assosiasjon) og med periferien (projeksjon), I.P. Pavlov skapte læren om den funksjonelle enheten til nevroner som tilhører forskjellige deler av nervesystemet - fra reseptorer i periferien til hjernebarken - læren om analysatorer. Det vi kaller senteret er den høyeste, kortikale delen av analysatoren. Hver analysator er knyttet til visse områder av hjernebarken (fig. 64).
I.P. Pavlov gjør betydelige justeringer av de tidligere ideene om de begrensede territoriene til kortikale sentre, til læren om den smale lokaliseringen av funksjoner. Dette er hva han sier om projeksjonen av reseptorer inn i hjernebarken.
"Hvert perifert reseptorapparat har i cortex et sentralt, spesielt, isolert territorium, som sin terminalstasjon, som representerer dens eksakte projeksjon. Her, takket være en spesiell design, kanskje et tettere arrangement av celler, flere celleforbindelser og fravær av celler med andre funksjoner, oppstår de mest komplekse irritasjonene, dannes (høyere syntese) og deres nøyaktige differensiering oppnås (høyere analyse) . Men disse reseptorelementene sprer seg videre over en veldig lang avstand, kanskje gjennom hele cortex.» Denne konklusjonen, basert på omfattende eksperimentelle og fysiologiske studier, er helt i samsvar med de siste morfologiske dataene om umuligheten av nøyaktig å avgrense kortikale cyto-arkitektoniske felt.
Følgelig kan ikke funksjonene til analysatorene (eller, med andre ord, arbeidet til det første signalsystemet) bare assosieres med de kortikale projeksjonssonene (kjernene til analysatorene). Dessuten er det umulig å begrense de mest komplekse, rent menneskelige funksjonene - funksjonene til det andre signalsystemet.
I.P. Pavlov definerer funksjonene til menneskelige signalsystemer som følger. "Jeg ser for meg hele komplekset av høyere nervøs aktivitet som dette. Hos høyerestående dyr, opp til og med mennesker, er den første instansen for organismens komplekse relasjoner med miljøet subcortex nærmest halvkulene med sine mest komplekse ubetingede reflekser (vår terminologi), instinkter, drifter, affekter, følelser (ulike , vanlig terminologi). Disse refleksene er forårsaket av relativt få ubetingede eksterne agenter. Derav den begrensede orienteringen i miljø og samtidig en svak tilpasning.
Den andre instansen er hjernehalvdelene... Her oppstår det ved hjelp av en betinget forbindelse (assosiasjon) nytt prinsipp aktivitet: signalering av noen få, ubetingede eksterne agenter av en utallig masse andre agenter, som stadig blir analysert og syntetisert, noe som gjør det mulig for en veldig stor orientering i samme miljø og samtidig en mye større tilpasning. Dette utgjør det eneste signalsystemet i dyrekroppen og det første hos mennesker.
Hos en person legges et annet signalsystem til, som signaliserer det første systemet - tale, dets basis eller basalkomponent - kinestetisk stimulering av taleorganene. Dette introduserer et nytt prinsipp for nervøs aktivitet - abstraksjon og sammen generalisering av utallige signaler fra det forrige systemet, i sin tur, igjen med analyse og syntese av disse første generaliserte signalene - et prinsipp som bestemmer grenseløs orientering i omverdenen og skaper den høyeste menneskelig tilpasning - vitenskap, både i form av en universell menneskelig empiri og i dens spesialiserte form."
Arbeidet til det andre signalsystemet er uløselig knyttet til funksjonene til alle analysatorer, derfor er det umulig å forestille seg lokaliseringen av de komplekse funksjonene til det andre signalsystemet i noen begrensede kortikale felt.
Betydningen av arven etterlatt oss av den store fysiologen for riktig utvikling av læren om lokalisering av funksjoner i hjernebarken er ekstremt stor. I.P. Pavlov la grunnlaget for en ny doktrine om dynamisk lokalisering av funksjoner i cortex. Konsepter om dynamisk lokalisering antyder muligheten for å bruke de samme kortikale strukturene i ulike kombinasjoner for å betjene ulike komplekse kortikale funksjoner.
Mens vi opprettholder en rekke klinisk etablerte definisjoner og tolkninger, vil vi prøve å gjøre noen justeringer av vår presentasjon i lys av læren til I.P. Pavlova om nervesystemet og dets patologi.
Så først av alt må vi vurdere spørsmålet om de såkalte projeksjons- og foreningssentrene. Den vanlige ideen om motoriske, sensoriske og andre projeksjonssentre (fremre og bakre sentrale gyri, visuelle, auditive sentre, etc.) er assosiert med konseptet om en ganske begrenset lokalisering i et gitt område av cortex av en eller en annen funksjon, og dette senteret er direkte relatert til de underliggende nevrale enhetene, og deretter med periferien, dens ledere (derav definisjonen - "projektiv"). Et eksempel på et slikt senter og dets leder er for eksempel den fremre sentrale gyrus og pyramidekanalen; fissura calcarina og radiatio optica, etc. Projeksjonssentre er forbundet gjennom assosiasjonsveier med andre sentre, med overflaten av cortex. Disse brede og kraftige assosiasjonsveiene bestemmer muligheten for kombinert aktivitet av ulike kortikale områder, etablering av nye forbindelser, og følgelig dannelsen av betingede reflekser.
"Associasjonssentre", i motsetning til projeksjonssentre, har ikke en direkte forbindelse med de underliggende delene av nervesystemet og periferien; de er bare forbundet med andre områder av cortex, inkludert "projeksjonssentre". Et eksempel på et «assosiasjonssenter» er det såkalte «senteret for stereognose» i parietallappen, lokalisert bak den bakre sentrale gyrus (fig. 65). Den bakre sentrale gyrus mottar gjennom de thalamo-kortikale banene individuelle irritasjoner som oppstår når hånden kjenner et objekt: taktil, form og størrelse (ledd-muskulær følelse), vekt, temperatur, etc. Alle disse følelsene overføres gjennom assosiasjonsfibre fra den bakre sentrale gyrusen til det "stereognostiske senteret", hvor de kombineres og skaper et felles sansebilde av objektet. Forbindelsene til det "stereognostiske senteret" med resten av cortex gjør det mulig å identifisere og sammenligne dette bildet med ideen om et gitt objekt, dets egenskaper, formål osv., allerede i minnet. (dvs. analyse og syntese av persepsjon utføres). Dette "senteret" har derfor ikke en direkte forbindelse med de underliggende delene av nervesystemet og er forbundet med assosiasjonsfibre med en rekke andre felt i hjernebarken.
Inndelingen av sentra i projeksjon og assosiasjon virker for oss feil. De store halvkulene er et sett med analysatorer for å analysere på den ene siden den ytre verden og på den andre interne prosesser. De perseptive sentrene i cortex ser ut til å være svært kompliserte og geografisk ekstremt utbredt. De øvre lagene i hjernebarken er faktisk helt okkupert av persepsjonssentre eller, i terminologien til I.P. Pavlov, "hjerneendene til analysatorer."
Fra alle lappene, fra de nedre lagene av cortex, er det efferente ledere som forbinder de kortikale endene av analysatorene med eksekutivorganene gjennom det subkortikale, stamme- og spinalapparatet. Et eksempel på en slik efferent leder er pyramidekanalen - dette interneuronet mellom den kinestetiske (motoriske) analysatoren og den perifere motorneuronen.
Hvordan kan vi så, fra dette synspunktet, forene posisjonen om tilstedeværelsen av motoriske projeksjonssentre (i den fremre sentrale gyrusen, øyerotasjonssenteret, etc.), når den er slått av, opplever en person lammelse, og når irritert, kramper med en helt klar somatotopisk fordeling og korrespondanse? Her snakker vi bare om skade på det motoriske projeksjonsområdet for de pyramidale kanalene, og ikke på "projeksjonsmotorsentrene."
Det er ingen tvil om at "frivillige" bevegelser er betingede motoriske reflekser, det vil si bevegelser som har utviklet seg, "tråkket ned" i prosessen med individuell livserfaring: men i utviklingen, organiseringen og den allerede skapte aktiviteten til skjelettmuskulaturen, avhenger alt av på den afferente enheten - hud- og motoranalysator (klinisk - hud- og ledd-muskulær følsomhet, mer bredt - kinestetisk sans), uten hvilken subtil og presis koordinering av en motorisk handling er umulig.
Ris. 64. Kortikale seksjoner av analysatorer (diagram).
a - ytre overflate; b - indre overflate. Rød - hudanalysator; gul - auditiv analysator: blå - visuell analysator; grønn - luktanalysator; stiplet linje - motoranalysator.
Den motoriske analysatoren (hvis oppgave er analyse og syntese av "frivillige" bevegelser) samsvarer ikke i det hele tatt med ideen om kortikale motoriske "projeksjons"-sentre med bestemte grenser for sistnevnte og en klar somatotopisk fordeling. Den motoriske analysatoren, som alle analysatorer, er forbundet med svært brede områder av cortex, og den motoriske funksjonen (i forhold til "frivillige" bevegelser) er ekstremt kompleks (hvis vi ikke bare tar hensyn til determinismen av bevegelser og atferd generelt , ikke bare kompleksiteten til handlingskomplekser, men også de afferente kinestetiske systemene, og orientering i forhold til miljøet og deler av ens egen kropp i rommet, etc.).
Hva kommer ideen om "projeksjonssentre" ned til? Det ble hevdet at sistnevnte representerte en slags inngang eller utgang "trigger gate" for impulser som kommer inn i cortex eller kommer fra den. Og hvis vi aksepterer at "motoriske projeksjonskortikale sentre" bare er slike "porter" (for det brede konseptet med en motoranalysator er absolutt assosiert med funksjonen til analyse og syntese), så bør vi anta at innenfor den fremre sentrale gyrus (og i territorier som ligner på den), og da bare i visse av lagene, er det et motorisk projeksjonsområde eller sone.
Hvordan kan vi da forestille oss de gjenværende "projeksjons"-sentrene (hudfølsomhet, syn, hørsel, smak, lukt) assosiert med andre (ikke-kinestetiske) afferente systemer? Vi tror at det ikke er noen grunnleggende forskjell her: faktisk, både i regionen av den bakre sentrale gyrus, og innenfor fissurae calcarinae, etc., strømmer impulser fra periferien til cellene i et bestemt lag av cortex, som er «projisert» her, og analyse og syntese skjer innenfor mange lag og brede områder.
Følgelig, i hver analysator (dens kortikale seksjon), inkludert den motoriske, er det et område eller sone som "projiserer" til periferien (motorisk område) eller som periferien "projiseres" inn i (sensitive områder, inkludert kinestetiske reseptorer for motoranalysatoren).
Det er akseptabelt at "projeksjonskjernen til analysatoren" kan identifiseres med begrepet en motorisk eller sensitiv projeksjonssone. Maksimal brudd, skrev I.I. Pavlov, analyse og syntese oppstår når nettopp en slik "projeksjonskjerne" er skadet; Hvis. Hvis vi tar den maksimale dysfunksjonen til analysatoren som en reell maksimal "sammenbrudd", som objektivt sett er helt korrekt, er den største manifestasjonen av skade på motoranalysatoren sentral lammelse, og den sensoriske er anestesi. Fra dette synspunktet ville det være riktig å identifisere konseptet "analysatorkjerne" med konseptet "projeksjonsområde til analysatoren".
Ris. 65. Tap av funksjoner observert med skade på ulike deler av hjernebarken (ytre overflate).
2 - visuelle forstyrrelser (hemianopsia); 3 - følsomhetsforstyrrelser; 4 - sentral lammelse eller parese; 5 - agraphia; 6 - kortikal lammelse av blikk og hoderotasjon i motsatt retning; 7 - motorisk afasi; 8 - hørselsforstyrrelser (ikke observert med ensidige lesjoner); 9 - amnestisk afasi; 10 - alexia; 11 - visuell agnosi (med bilateral skade); 12 - astereognosi; 13 - apraxia; 14 - sensorisk afasi.
Basert på ovenstående anser vi det som riktig å erstatte konseptet med et projeksjonssenter med konseptet om et projeksjonsområde i analysatorsonen. Da er inndelingen av kortikale "sentre" i projeksjon og assosiasjon ubegrunnet: det er analysatorer (deres kortikale seksjoner) og, innenfor deres grenser, projeksjonsområder.
Forelesning 12. LOKALISERING AV FUNKSJONER I DE STORE HELVEKSJONER CORTEX Kortikale soner. Projeksjon kortikale soner: primær og sekundær. Motoriske (motoriske) soner i hjernebarken. Tertiære kortikale soner.
 Tap av funksjoner observert med skade på ulike deler av cortex (den indre overflaten). 1 - luktforstyrrelser (ikke observert med ensidige lesjoner); 2 - visuelle forstyrrelser (hemianopsia); 3 - følsomhetsforstyrrelser; 4 - sentral lammelse eller parese. Data eksperimentell forskning ved ødeleggelse eller fjerning av visse områder av cortex og kliniske observasjoner indikerer at funksjoner er begrenset til aktiviteten til visse områder av cortex. Et område av hjernebarken som har en spesifikk funksjon kalles den kortikale sonen. Det er projeksjon, assosiative kortikale soner og motoriske (motoriske) soner.
Tap av funksjoner observert med skade på ulike deler av cortex (den indre overflaten). 1 - luktforstyrrelser (ikke observert med ensidige lesjoner); 2 - visuelle forstyrrelser (hemianopsia); 3 - følsomhetsforstyrrelser; 4 - sentral lammelse eller parese. Data eksperimentell forskning ved ødeleggelse eller fjerning av visse områder av cortex og kliniske observasjoner indikerer at funksjoner er begrenset til aktiviteten til visse områder av cortex. Et område av hjernebarken som har en spesifikk funksjon kalles den kortikale sonen. Det er projeksjon, assosiative kortikale soner og motoriske (motoriske) soner.
 Den projeksjonskortikale sonen er den kortikale representasjonen av analysatoren. Nevroner av projeksjonssoner mottar signaler av en modalitet (visuell, auditiv, etc.). Det er: - primære projeksjonssoner; - sekundære projeksjonssoner, som gir en integrerende funksjon av persepsjon. I sonen til en bestemt analysator skilles også tertiære felt eller assosiative soner.
Den projeksjonskortikale sonen er den kortikale representasjonen av analysatoren. Nevroner av projeksjonssoner mottar signaler av en modalitet (visuell, auditiv, etc.). Det er: - primære projeksjonssoner; - sekundære projeksjonssoner, som gir en integrerende funksjon av persepsjon. I sonen til en bestemt analysator skilles også tertiære felt eller assosiative soner.
 De primære projeksjonsfeltene til cortex mottar informasjon mediert gjennom det minste antallet brytere i subcortex (thalamus, diencephalon). Overflaten til perifere reseptorer projiseres så å si på disse feltene. Nervefibre kommer inn i hjernebarken hovedsakelig fra thalamus (disse er afferente innganger).
De primære projeksjonsfeltene til cortex mottar informasjon mediert gjennom det minste antallet brytere i subcortex (thalamus, diencephalon). Overflaten til perifere reseptorer projiseres så å si på disse feltene. Nervefibre kommer inn i hjernebarken hovedsakelig fra thalamus (disse er afferente innganger).
 Projeksjonssonene til analysesystemene okkuperer den ytre overflaten av hjernens bakre cortex. Dette inkluderer de visuelle (occipitale), auditive (temporale) og sensoriske (parietale) områdene i cortex. Den kortikale avdelingen inkluderer også representasjon av smak, lukt, visceral følsomhet
Projeksjonssonene til analysesystemene okkuperer den ytre overflaten av hjernens bakre cortex. Dette inkluderer de visuelle (occipitale), auditive (temporale) og sensoriske (parietale) områdene i cortex. Den kortikale avdelingen inkluderer også representasjon av smak, lukt, visceral følsomhet
 Primære sensoriske områder (Brodmann-områder): visuelle - 17, auditive - 41 og somatosensoriske - 1, 2, 3 (samlet kalles de sensorisk cortex), motorisk (4) og premotorisk (6) cortex
Primære sensoriske områder (Brodmann-områder): visuelle - 17, auditive - 41 og somatosensoriske - 1, 2, 3 (samlet kalles de sensorisk cortex), motorisk (4) og premotorisk (6) cortex
 Primære sensoriske områder (Brodmann-områder): visuelle - 17, auditive - 41 og somatosensoriske - 1, 2, 3 (samlet kalles de sensorisk cortex), motorisk (4) og premotorisk (6) cortex Hvert felt i hjernebarken er karakterisert av en spesiell sammensetning nevroner, deres plassering og forbindelser mellom dem. Feltene i sansebarken, der den primære behandlingen av informasjon fra sanseorganer skjer, skiller seg sterkt fra den primære motoriske cortex, som er ansvarlig for å generere kommandoer for frivillige muskelbevegelser.
Primære sensoriske områder (Brodmann-områder): visuelle - 17, auditive - 41 og somatosensoriske - 1, 2, 3 (samlet kalles de sensorisk cortex), motorisk (4) og premotorisk (6) cortex Hvert felt i hjernebarken er karakterisert av en spesiell sammensetning nevroner, deres plassering og forbindelser mellom dem. Feltene i sansebarken, der den primære behandlingen av informasjon fra sanseorganer skjer, skiller seg sterkt fra den primære motoriske cortex, som er ansvarlig for å generere kommandoer for frivillige muskelbevegelser.
 I den motoriske cortex dominerer nevroner formet som pyramider, og sensorisk cortex er hovedsakelig representert av nevroner hvis kroppsform ligner korn eller granulat, og det er derfor de kalles granulære. Struktur av hjernebarken I. molekylær II. ekstern granulær III. ytre pyramideformet IV. indre granulær V. ganglion (gigantiske pyramider) VI. polymorf
I den motoriske cortex dominerer nevroner formet som pyramider, og sensorisk cortex er hovedsakelig representert av nevroner hvis kroppsform ligner korn eller granulat, og det er derfor de kalles granulære. Struktur av hjernebarken I. molekylær II. ekstern granulær III. ytre pyramideformet IV. indre granulær V. ganglion (gigantiske pyramider) VI. polymorf
 Nevroner i de primære projeksjonssonene i cortex har generelt høyest spesifisitet. For eksempel reagerer nevroner i de visuelle områdene selektivt på fargenyanser, bevegelsesretning, linjers karakter osv. Men i primærsonene til individuelle områder av cortex er det også multimodale type nevroner som reagerer på flere typer stimuli og nevroner hvis reaksjon reflekterer påvirkningen av uspesifikke (limbicoretikulære) systemer.
Nevroner i de primære projeksjonssonene i cortex har generelt høyest spesifisitet. For eksempel reagerer nevroner i de visuelle områdene selektivt på fargenyanser, bevegelsesretning, linjers karakter osv. Men i primærsonene til individuelle områder av cortex er det også multimodale type nevroner som reagerer på flere typer stimuli og nevroner hvis reaksjon reflekterer påvirkningen av uspesifikke (limbicoretikulære) systemer.
 Projeksjon afferente fibre ender i primærfeltene. Dermed er felt 1 og 3, som okkuperer de mediale og laterale overflatene til den bakre sentrale gyrusen, de primære projeksjonsfeltene for kutan følsomhet av kroppsoverflaten.
Projeksjon afferente fibre ender i primærfeltene. Dermed er felt 1 og 3, som okkuperer de mediale og laterale overflatene til den bakre sentrale gyrusen, de primære projeksjonsfeltene for kutan følsomhet av kroppsoverflaten.
 Den funksjonelle organiseringen av projeksjonssoner i cortex er basert på prinsippet om lokal lokalisering. Perceptive elementer som ligger ved siden av hverandre i periferien (for eksempel områder av huden) projiseres på den kortikale overflaten også ved siden av hverandre.
Den funksjonelle organiseringen av projeksjonssoner i cortex er basert på prinsippet om lokal lokalisering. Perceptive elementer som ligger ved siden av hverandre i periferien (for eksempel områder av huden) projiseres på den kortikale overflaten også ved siden av hverandre.
 De nedre lemmene er representert i den mediale delen, og projeksjoner av reseptorfeltene på hudens overflate av hodet er plassert lavest på den laterale delen av gyrusen. I dette tilfellet projiseres områder av kroppsoverflaten rikt utstyrt med reseptorer (fingre, lepper, tunge) på stort område cortex enn områder med færre reseptorer (lår, rygg, skulder).
De nedre lemmene er representert i den mediale delen, og projeksjoner av reseptorfeltene på hudens overflate av hodet er plassert lavest på den laterale delen av gyrusen. I dette tilfellet projiseres områder av kroppsoverflaten rikt utstyrt med reseptorer (fingre, lepper, tunge) på stort område cortex enn områder med færre reseptorer (lår, rygg, skulder).

 Feltene 17-19, som ligger i den occipitale lappen, er det visuelle sentrum av cortex; felt 17, som opptar selve occipitalpolen, er primært. 18. og 19. felt ved siden av det utfører funksjonen som sekundære felt og mottar inndata fra 17. felt.
Feltene 17-19, som ligger i den occipitale lappen, er det visuelle sentrum av cortex; felt 17, som opptar selve occipitalpolen, er primært. 18. og 19. felt ved siden av det utfører funksjonen som sekundære felt og mottar inndata fra 17. felt.
 De auditive projeksjonsfeltene er lokalisert i tinninglappene (41, 42). Ved siden av dem, på grensen til tinning-, occipital- og parietallappene, ligger den 37., 39. og 40., kun karakteristisk for den menneskelige hjernebarken. For de fleste inneholder disse feltene på venstre hjernehalvdel talesenteret, som er ansvarlig for oppfatningen av muntlig og skriftlig tale.
De auditive projeksjonsfeltene er lokalisert i tinninglappene (41, 42). Ved siden av dem, på grensen til tinning-, occipital- og parietallappene, ligger den 37., 39. og 40., kun karakteristisk for den menneskelige hjernebarken. For de fleste inneholder disse feltene på venstre hjernehalvdel talesenteret, som er ansvarlig for oppfatningen av muntlig og skriftlig tale.

 Sekundære projeksjonsfelt, som mottar informasjon fra de primære, er plassert ved siden av dem. Nevronene i disse feltene er preget av oppfatningen av komplekse tegn på stimuli, men samtidig bevares spesifisiteten som tilsvarer nevronene i primærsonene. Komplikasjonen av detektoregenskapene til nevroner i sekundærsonene kan oppstå gjennom konvergens av nevroner i primærsonene på dem. I sekundærsonene (18. og 19. Brodmann-felt) vises detektorer mer komplekse elementer kontur: kanter med begrensede linjelengder, hjørner med forskjellige orienteringer, etc.
Sekundære projeksjonsfelt, som mottar informasjon fra de primære, er plassert ved siden av dem. Nevronene i disse feltene er preget av oppfatningen av komplekse tegn på stimuli, men samtidig bevares spesifisiteten som tilsvarer nevronene i primærsonene. Komplikasjonen av detektoregenskapene til nevroner i sekundærsonene kan oppstå gjennom konvergens av nevroner i primærsonene på dem. I sekundærsonene (18. og 19. Brodmann-felt) vises detektorer mer komplekse elementer kontur: kanter med begrensede linjelengder, hjørner med forskjellige orienteringer, etc.
 Motoriske (motoriske) soner i hjernebarken er områder av den motoriske cortex, hvis nevroner forårsaker en motorisk handling. De motoriske områdene av cortex er lokalisert i den presentrale gyrusen til frontallappen (foran projeksjonssonene med kutan følsomhet). Denne delen av cortex er okkupert av felt 4 og 6. Fra V-laget til disse feltene utgår pyramidekanalen, som ender på de motoriske nevronene i ryggmargen.
Motoriske (motoriske) soner i hjernebarken er områder av den motoriske cortex, hvis nevroner forårsaker en motorisk handling. De motoriske områdene av cortex er lokalisert i den presentrale gyrusen til frontallappen (foran projeksjonssonene med kutan følsomhet). Denne delen av cortex er okkupert av felt 4 og 6. Fra V-laget til disse feltene utgår pyramidekanalen, som ender på de motoriske nevronene i ryggmargen.
 Premotorisk sone (felt 6) Den premotoriske sonen i cortex er plassert foran den motoriske sonen, den er ansvarlig for muskeltonus og koordinerte bevegelser av hodet og overkroppen. De viktigste efferente utgangene fra cortex er aksonene til lag V-pyramider. Dette er efferente, motoriske nevroner involvert i reguleringen av motoriske funksjoner.
Premotorisk sone (felt 6) Den premotoriske sonen i cortex er plassert foran den motoriske sonen, den er ansvarlig for muskeltonus og koordinerte bevegelser av hodet og overkroppen. De viktigste efferente utgangene fra cortex er aksonene til lag V-pyramider. Dette er efferente, motoriske nevroner involvert i reguleringen av motoriske funksjoner.


 Tertiære eller interanalyzersoner (assosiativ) Prefrontal sone (felt 9, 10, 45, 46, 47, 11), parietotemporal (felt 39, 40) Afferente og efferente projeksjonssoner av cortex opptar et relativt lite område. Det meste av overflaten av cortex er okkupert av tertiære eller interanalyzersoner, kalt assosiative soner. De mottar multimodale input fra de sensoriske områdene i cortex og thalamus assosiative kjerner og har utganger til de motoriske områdene i cortex. Associative soner gir integrering av sensoriske input og spiller en betydelig rolle i mental aktivitet (læring, tenkning).
Tertiære eller interanalyzersoner (assosiativ) Prefrontal sone (felt 9, 10, 45, 46, 47, 11), parietotemporal (felt 39, 40) Afferente og efferente projeksjonssoner av cortex opptar et relativt lite område. Det meste av overflaten av cortex er okkupert av tertiære eller interanalyzersoner, kalt assosiative soner. De mottar multimodale input fra de sensoriske områdene i cortex og thalamus assosiative kjerner og har utganger til de motoriske områdene i cortex. Associative soner gir integrering av sensoriske input og spiller en betydelig rolle i mental aktivitet (læring, tenkning).
 Funksjoner til ulike områder av neocortex: 5 3 7 6 4 1 2 Hukommelse, behov Utløsende atferd 1. Occipital lobe - visuell cortex. 2. Temporallappen – auditiv cortex. 3. Fremre del av parietallappen – smerte, hud- og muskelfølsomhet. 4. Inne i lateral sulcus (insula) – vestibulær følsomhet og smak. 5. Den bakre delen av frontallappen er den motoriske cortex. 6. Den bakre delen av parietallappene og tinninglappene er den assosiative parietale cortex: den kombinerer signalstrømmer fra forskjellige sansesystemer, talesentre og tenkesentre. 7. Fremre del av frontallappen - assosiativ frontal cortex: tar i betraktning sensoriske signaler, signaler fra sentrene for behov, hukommelse og tenkning, tar beslutninger om lansering av atferdsprogrammer ("senter for vilje og initiativ").
Funksjoner til ulike områder av neocortex: 5 3 7 6 4 1 2 Hukommelse, behov Utløsende atferd 1. Occipital lobe - visuell cortex. 2. Temporallappen – auditiv cortex. 3. Fremre del av parietallappen – smerte, hud- og muskelfølsomhet. 4. Inne i lateral sulcus (insula) – vestibulær følsomhet og smak. 5. Den bakre delen av frontallappen er den motoriske cortex. 6. Den bakre delen av parietallappene og tinninglappene er den assosiative parietale cortex: den kombinerer signalstrømmer fra forskjellige sansesystemer, talesentre og tenkesentre. 7. Fremre del av frontallappen - assosiativ frontal cortex: tar i betraktning sensoriske signaler, signaler fra sentrene for behov, hukommelse og tenkning, tar beslutninger om lansering av atferdsprogrammer ("senter for vilje og initiativ").
 Enkelte store assosiasjonsområder er plassert ved siden av de tilsvarende sanseområdene. Noen assosiative soner utfører kun en begrenset spesialisert funksjon og er koblet til andre assosiative sentre som er i stand til å utsette informasjon for videre behandling. For eksempel analyserer det auditive assosiasjonsområdet lyder, kategoriserer dem, og sender deretter signaler til mer spesialiserte områder, som for eksempel taleforeningsområdet, der betydningen av ord som høres oppfattes.
Enkelte store assosiasjonsområder er plassert ved siden av de tilsvarende sanseområdene. Noen assosiative soner utfører kun en begrenset spesialisert funksjon og er koblet til andre assosiative sentre som er i stand til å utsette informasjon for videre behandling. For eksempel analyserer det auditive assosiasjonsområdet lyder, kategoriserer dem, og sender deretter signaler til mer spesialiserte områder, som for eksempel taleforeningsområdet, der betydningen av ord som høres oppfattes.
 Assosiasjonsfeltene til parietallappen kombinerer informasjon som kommer fra den somatosensoriske cortex (fra hud, muskler, sener og ledd angående kroppsposisjon og bevegelse) med visuell og auditiv informasjon som kommer fra de visuelle og auditive cortexene i occipital- og temporallappene. Denne kombinerte informasjonen hjelper deg med å få en nøyaktig forståelse av din egen kropp mens du beveger deg rundt i miljøet.
Assosiasjonsfeltene til parietallappen kombinerer informasjon som kommer fra den somatosensoriske cortex (fra hud, muskler, sener og ledd angående kroppsposisjon og bevegelse) med visuell og auditiv informasjon som kommer fra de visuelle og auditive cortexene i occipital- og temporallappene. Denne kombinerte informasjonen hjelper deg med å få en nøyaktig forståelse av din egen kropp mens du beveger deg rundt i miljøet.
 Wernickes område og Brocas område er to områder av hjernen som er involvert i prosessen med å reprodusere og forstå informasjon relatert til tale. Begge områdene ligger langs den sylviske sprekken (den laterale sprekken i hjernehalvdelene). Afasi er et fullstendig eller delvis tap av tale forårsaket av lokale lesjoner i hjernen.
Wernickes område og Brocas område er to områder av hjernen som er involvert i prosessen med å reprodusere og forstå informasjon relatert til tale. Begge områdene ligger langs den sylviske sprekken (den laterale sprekken i hjernehalvdelene). Afasi er et fullstendig eller delvis tap av tale forårsaket av lokale lesjoner i hjernen.






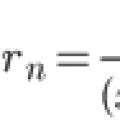 Taylor-serieutvidelse Omtrentlig løsning av Cauchy-problemet for den vanlige
Taylor-serieutvidelse Omtrentlig løsning av Cauchy-problemet for den vanlige Hva læres IKKE på skolen Hva læres ikke på skolen spør
Hva læres IKKE på skolen Hva læres ikke på skolen spør Money Thinking Formel (A
Money Thinking Formel (A