Vekst og reproduksjon av bakterier. Reproduksjonsfaser
 Veksten av en bakteriecelle skal forstås som en økning i massen av dens cytoplasma, som oppstår som et resultat av syntesen av cellulært materiale under ernæring. Veksten til en bakteriepopulasjon går gjennom 4 stadier: 1) lagfase, 2) eksponentiell eller logaritmisk fase, 3) stasjonær fase, 4) døende fase.
Veksten av en bakteriecelle skal forstås som en økning i massen av dens cytoplasma, som oppstår som et resultat av syntesen av cellulært materiale under ernæring. Veksten til en bakteriepopulasjon går gjennom 4 stadier: 1) lagfase, 2) eksponentiell eller logaritmisk fase, 3) stasjonær fase, 4) døende fase.
 LAG-FASE (4-5 timer) Oppstår etter at frøet er introdusert i mediet. Dette er perioden for tilpasning av bakterier til næringsmediet, når differensiell aktivering av ekso- og endoenzymer skjer for den påfølgende implementeringen av enzym-substratreaksjonen. Med stabilt DNA-innhold er det en kraftig økning i bakterieprotein og RNA.
LAG-FASE (4-5 timer) Oppstår etter at frøet er introdusert i mediet. Dette er perioden for tilpasning av bakterier til næringsmediet, når differensiell aktivering av ekso- og endoenzymer skjer for den påfølgende implementeringen av enzym-substratreaksjonen. Med stabilt DNA-innhold er det en kraftig økning i bakterieprotein og RNA.
 LAG FASE (4-5 timer) Varigheten av etterslepfasen er vanligvis kort, målt i timer og avhenger av typen bakterie, mangfoldet av inokulering på et gitt medium, tilstanden til kulturen, temperaturen som brukes til dyrking, og sammensetningen av næringsmediet. I fravær av synlige tegn på vekst i lagfasen, oppstår en økning i biomasse, som et resultat av at størrelsen på bakteriecellen øker flere ganger.
LAG FASE (4-5 timer) Varigheten av etterslepfasen er vanligvis kort, målt i timer og avhenger av typen bakterie, mangfoldet av inokulering på et gitt medium, tilstanden til kulturen, temperaturen som brukes til dyrking, og sammensetningen av næringsmediet. I fravær av synlige tegn på vekst i lagfasen, oppstår en økning i biomasse, som et resultat av at størrelsen på bakteriecellen øker flere ganger.
 LAG FASE (4 -5 timer) Etter å ha nådd en viss størrelse, har "akkumulert" den nødvendige mengden protein, RNA og DNA, aktivering av ekso- og endoenzymer, begynner bakteriecellen å dele seg aktivt. Bakterier formerer seg ved tverrgående celledeling.
LAG FASE (4 -5 timer) Etter å ha nådd en viss størrelse, har "akkumulert" den nødvendige mengden protein, RNA og DNA, aktivering av ekso- og endoenzymer, begynner bakteriecellen å dele seg aktivt. Bakterier formerer seg ved tverrgående celledeling.
 LOGARITMISK VEKSTFASE (5 - 6 timer) Dette er reproduksjonsfasen, utført gjennom den binære deling av modercellen i to datterceller. "Kjedereaksjonen med progressivt akselererende binær fisjon av bakterieceller fører til en rask økning i bakteriemasse i næringsmediet, intensivt forbruk av energisubstratet og akkumulering av bakterielle metabolske produkter.
LOGARITMISK VEKSTFASE (5 - 6 timer) Dette er reproduksjonsfasen, utført gjennom den binære deling av modercellen i to datterceller. "Kjedereaksjonen med progressivt akselererende binær fisjon av bakterieceller fører til en rask økning i bakteriemasse i næringsmediet, intensivt forbruk av energisubstratet og akkumulering av bakterielle metabolske produkter.
 STASJONÆR VEKSTFASE Som et resultat blir miljøet stadig mer ugunstig for videre vekst og reproduksjon av bakterier. Under den stasjonære fasen forblir reproduksjonshastigheten konstant. Avhengig av hvilken type bakterier som dyrkes, kan den vare i lang tid, hvoretter det fjerde stadiet oppstår -
STASJONÆR VEKSTFASE Som et resultat blir miljøet stadig mer ugunstig for videre vekst og reproduksjon av bakterier. Under den stasjonære fasen forblir reproduksjonshastigheten konstant. Avhengig av hvilken type bakterier som dyrkes, kan den vare i lang tid, hvoretter det fjerde stadiet oppstår -
 DØDEFASE Den døende fasen er preget av den progressive døden av bakterieceller på en logaritmisk måte. Varigheten av denne fasen varierer fra 48 timer til flere uker.
DØDEFASE Den døende fasen er preget av den progressive døden av bakterieceller på en logaritmisk måte. Varigheten av denne fasen varierer fra 48 timer til flere uker.
 Naturen til bakterievekst på flytende næringsmedier er forskjellig - diffus turbiditet i næringsmediet, - dannelse av en film eller sediment (bunnvekst), - vekst i form av en "bomullskule". Vekstmønsteret på flytende næringsmedium brukes til å differensiere bakterier.
Naturen til bakterievekst på flytende næringsmedier er forskjellig - diffus turbiditet i næringsmediet, - dannelse av en film eller sediment (bunnvekst), - vekst i form av en "bomullskule". Vekstmønsteret på flytende næringsmedium brukes til å differensiere bakterier.
 Næringsmedier For dyrking av bakterier under laboratorieforhold brukes kunstige næringsmedier av ulike sammensetninger. Konvensjonelle eller enkle næringsmedier (kjøttpeptonagar, kjøttpeptonbuljong) brukes til innledende avlinger (primær). Komplekse medier inkluderer selektive og differensialdiagnostiske næringsmedier.
Næringsmedier For dyrking av bakterier under laboratorieforhold brukes kunstige næringsmedier av ulike sammensetninger. Konvensjonelle eller enkle næringsmedier (kjøttpeptonagar, kjøttpeptonbuljong) brukes til innledende avlinger (primær). Komplekse medier inkluderer selektive og differensialdiagnostiske næringsmedier.
 Næringsmedier Elektive medier sikrer vekst av kun en viss type mikroorganismer, mens den medfølgende mikrofloraen undertrykkes av spesielle tilsetningsstoffer. Differensialdiagnostiske næringsmedier brukes til å studere de biokjemiske egenskapene til mikroorganismer og gjøre det mulig å differensiere bakterier ved enzymatisk aktivitet.
Næringsmedier Elektive medier sikrer vekst av kun en viss type mikroorganismer, mens den medfølgende mikrofloraen undertrykkes av spesielle tilsetningsstoffer. Differensialdiagnostiske næringsmedier brukes til å studere de biokjemiske egenskapene til mikroorganismer og gjøre det mulig å differensiere bakterier ved enzymatisk aktivitet.
 KLASSIFISERING AV MIKROORGANISMER Etter hvert som nye bakteriearter ble studert og identifisert, reflekterte hver nyopprettede klassifikasjon utviklingsnivået til vitenskapen. Klassifiseringen av mikroorganismer, det vil si systematiseringen av alle kjente arter, var basert på en rekke egenskaper:
KLASSIFISERING AV MIKROORGANISMER Etter hvert som nye bakteriearter ble studert og identifisert, reflekterte hver nyopprettede klassifikasjon utviklingsnivået til vitenskapen. Klassifiseringen av mikroorganismer, det vil si systematiseringen av alle kjente arter, var basert på en rekke egenskaper:

 Sekvens for å bestemme en mikroorganisme I. Hvilket rike tilhører den - prokaryoter eller eukaryoter II. Hvilken av hovedkategoriene tilhører den: 1. Gramnegative eubakterier som har cellevegger. 2. Gram-positive eubakterier med cellevegger. 3. Eubakterier som mangler cellevegger. 4. Arkebakterier.
Sekvens for å bestemme en mikroorganisme I. Hvilket rike tilhører den - prokaryoter eller eukaryoter II. Hvilken av hovedkategoriene tilhører den: 1. Gramnegative eubakterier som har cellevegger. 2. Gram-positive eubakterier med cellevegger. 3. Eubakterier som mangler cellevegger. 4. Arkebakterier.
 Totalt 35 grupper av mikroorganismer er kjent III. Hvilken gruppe innenfor 4 kategorier tilhører mikroorganismen: 1. Spiroketter 2. Aerobe /mikroaerofile, bevegelige, spiralformede/, vibroide, gramnegative bakterier. 3. Ikke-bevegelige gramnegative, buede bakterier. 4. Gram-negative, anaerobe, mikroaerofile staver og kokker.
Totalt 35 grupper av mikroorganismer er kjent III. Hvilken gruppe innenfor 4 kategorier tilhører mikroorganismen: 1. Spiroketter 2. Aerobe /mikroaerofile, bevegelige, spiralformede/, vibroide, gramnegative bakterier. 3. Ikke-bevegelige gramnegative, buede bakterier. 4. Gram-negative, anaerobe, mikroaerofile staver og kokker.
 I. gramnegative eubakterier med cellevegg 5. Fakultative anaerobe, gramnegative staver. 6. Gramnegative, anaerobe, rette, buede og spiralformede stenger. 7. Bakterier som utfører dissimilerende reduksjon av sulfat eller svovel. 8. Anaerobe gramnegative kokker. 9. Rickettsia og klamydia.
I. gramnegative eubakterier med cellevegg 5. Fakultative anaerobe, gramnegative staver. 6. Gramnegative, anaerobe, rette, buede og spiralformede stenger. 7. Bakterier som utfører dissimilerende reduksjon av sulfat eller svovel. 8. Anaerobe gramnegative kokker. 9. Rickettsia og klamydia.
 I. gramnegative eubakterier med cellevegg 10. Anoksygene fototrofe bakterier. 11. Oksygene fototrofe bakterier. 12. Aerobe kjemolitotrofe bakterier. 13. Spirende og/eller utvekstdannende bakterier. 14. Bakterier med deksler. 15. Ikke-fotosyntetiske glidebakterier som ikke danner fruktlegemer. 16. Glidende bakterier som danner fruktlegemer.
I. gramnegative eubakterier med cellevegg 10. Anoksygene fototrofe bakterier. 11. Oksygene fototrofe bakterier. 12. Aerobe kjemolitotrofe bakterier. 13. Spirende og/eller utvekstdannende bakterier. 14. Bakterier med deksler. 15. Ikke-fotosyntetiske glidebakterier som ikke danner fruktlegemer. 16. Glidende bakterier som danner fruktlegemer.
 II. Gram-positive eubakterier med cellevegger. 1. Gram-positive kokker. 2. Gram-positive staver og kokker som danner endosporer. 3. Gram-positive staver som ikke danner sporer og har regelmessig form. 4. Gram-positive staver med uregelmessig form som ikke danner sporer. 5. Mykobakterier. 6. Actinomycetes.
II. Gram-positive eubakterier med cellevegger. 1. Gram-positive kokker. 2. Gram-positive staver og kokker som danner endosporer. 3. Gram-positive staver som ikke danner sporer og har regelmessig form. 4. Gram-positive staver med uregelmessig form som ikke danner sporer. 5. Mykobakterier. 6. Actinomycetes.

 IV. Arkebakterier. 1. Metanogener. 2. Sulfatreduserende arkea. 3. Ekstremt halofile arkebakterier. 4. Arkebakterier som mangler cellevegg. 5. Ekstreme termofile og hypertermofiler som metaboliserer S
IV. Arkebakterier. 1. Metanogener. 2. Sulfatreduserende arkea. 3. Ekstremt halofile arkebakterier. 4. Arkebakterier som mangler cellevegg. 5. Ekstreme termofile og hypertermofiler som metaboliserer S
 Sekvens for å identifisere en mikroorganisme IV. Hvilken slekt tilhører mikroorganismen? V. Hvilken familie tilhører mikroorganismen? VI. Hvilken type mikroorganisme er det?
Sekvens for å identifisere en mikroorganisme IV. Hvilken slekt tilhører mikroorganismen? V. Hvilken familie tilhører mikroorganismen? VI. Hvilken type mikroorganisme er det?
 Konstruksjon av det taksonomiske navnet på mikroorganismen. 1. RIKE 2. KATEGORI. 3. GRUPPE. 4. STANG. 5. FAMILIE. 6. VISNING
Konstruksjon av det taksonomiske navnet på mikroorganismen. 1. RIKE 2. KATEGORI. 3. GRUPPE. 4. STANG. 5. FAMILIE. 6. VISNING
 Fordeler med den moderne klassifiseringen av mikroorganismer Den fylogenetiske systematiseringen som er opprettet til dags dato har alle fordelene og ulempene ved en klassifisering basert på en egenskap. Fordelene inkluderer den nesten fullstendige identiteten til resultatene oppnådd i forskjellige laboratorier rundt om i verden. For å etablere artsidentitet begynte de også å i tillegg evaluere graden av DNA-DNA-homologi ved å bruke typestammer.
Fordeler med den moderne klassifiseringen av mikroorganismer Den fylogenetiske systematiseringen som er opprettet til dags dato har alle fordelene og ulempene ved en klassifisering basert på en egenskap. Fordelene inkluderer den nesten fullstendige identiteten til resultatene oppnådd i forskjellige laboratorier rundt om i verden. For å etablere artsidentitet begynte de også å i tillegg evaluere graden av DNA-DNA-homologi ved å bruke typestammer.
 Ulemper ved den eksisterende klassifiseringen av mikroorganismer. Ulempen med den eksisterende klassifiseringen er at den ikke gir en ide om funksjonene til bakterier. Derfor er opprettelsen av en fenotypisk eller funksjonell klassifisering nå av stor betydning for praktiske mikrobiologer. For raskt å bestemme den taksonomiske posisjonen til mikroorganismer, bruk Bergey-determinanten. Denne referansepublikasjonen oppdateres kontinuerlig med nye grupper av isolater og trykkes med jevne mellomrom. Nå er den 11. utgaven aktuell.
Ulemper ved den eksisterende klassifiseringen av mikroorganismer. Ulempen med den eksisterende klassifiseringen er at den ikke gir en ide om funksjonene til bakterier. Derfor er opprettelsen av en fenotypisk eller funksjonell klassifisering nå av stor betydning for praktiske mikrobiologer. For raskt å bestemme den taksonomiske posisjonen til mikroorganismer, bruk Bergey-determinanten. Denne referansepublikasjonen oppdateres kontinuerlig med nye grupper av isolater og trykkes med jevne mellomrom. Nå er den 11. utgaven aktuell.
 Dannelse av en moderne klassifisering av mikroorganismer. På det nåværende stadiet utvikles identifisering av den fylogenetiske posisjonen til prokaryoter, inkludert ukultiverte, på grunnlag av nukleotidsekvenser av 16 S r RNA. Forbedret sekvensering og databehandlingsteknikker har gjort denne tilnærmingen praktisk talt ikke noe alternativ til å bestemme slekten til nye organismer. Beskrivelsen av nye bakterielle taxaer har skjedd i et veldig raskt tempo de siste 50 årene, takket være fremskritt i studiet av anaerober.
Dannelse av en moderne klassifisering av mikroorganismer. På det nåværende stadiet utvikles identifisering av den fylogenetiske posisjonen til prokaryoter, inkludert ukultiverte, på grunnlag av nukleotidsekvenser av 16 S r RNA. Forbedret sekvensering og databehandlingsteknikker har gjort denne tilnærmingen praktisk talt ikke noe alternativ til å bestemme slekten til nye organismer. Beskrivelsen av nye bakterielle taxaer har skjedd i et veldig raskt tempo de siste 50 årene, takket være fremskritt i studiet av anaerober.
 Forskjellen mellom klassifisering og identifikasjon I tillegg til klassifikasjoner finnes det i mikrobiologi ordninger for å identifisere isolerte bakteriekulturer. For å bygge et identifiseringsskjema, velg egenskaper ved mikroorganismer som er enkle å bestemme, og for klassifisering krever ofte bruk av komplekse metoder. I dette tilfellet bør identifikasjonsskjemaet inkludere et lite antall kjennetegn, og for taksonomisk bestemmelse bruker klassifiseringen så mange kjennetegn som mulig.
Forskjellen mellom klassifisering og identifikasjon I tillegg til klassifikasjoner finnes det i mikrobiologi ordninger for å identifisere isolerte bakteriekulturer. For å bygge et identifiseringsskjema, velg egenskaper ved mikroorganismer som er enkle å bestemme, og for klassifisering krever ofte bruk av komplekse metoder. I dette tilfellet bør identifikasjonsskjemaet inkludere et lite antall kjennetegn, og for taksonomisk bestemmelse bruker klassifiseringen så mange kjennetegn som mulig.
 TAKK FOR DIN OPPMERKSOMHET. DU HAR HØRT FOREDRAG nr. 3 OM MIKROBIOLOGI OM EMNET: «VEKST OG REPRODUKSJON AV MIKROORGANISMER. EVOLUSJON OG KLASSIFISERING AV MIKROORGANISMER".
TAKK FOR DIN OPPMERKSOMHET. DU HAR HØRT FOREDRAG nr. 3 OM MIKROBIOLOGI OM EMNET: «VEKST OG REPRODUKSJON AV MIKROORGANISMER. EVOLUSJON OG KLASSIFISERING AV MIKROORGANISMER".
Vekst og reproduksjon av bakterier. Mekanisme og reproduksjonshastighet. Faser av mikrobiell reproduksjon.
| Parameternavn | Betydning |
| Artikkel emne: | Vekst og reproduksjon av bakterier. Mekanisme og reproduksjonshastighet. Faser av mikrobiell reproduksjon. |
| Rubrikk (tematisk kategori) | Kultur |
1. Begreper om bakterievekst og reproduksjon
2. Bakteriepopulasjon
3. Kolonier
1 . For mikrobiologisk diagnostikk, studie av mikroorganismer og til bioteknologiske formål mikroorganismer dyrkes på kunstige næringsmedier.
Under bakterievekst
forstå økning i cellemasse uten å endre antallet i populasjonen som et resultat av koordinert reproduksjon av alle cellulære komponenter og strukturer.
Lagt ut på ref.rf
Øke antall celler i en populasjon av mikroorganismer betegnet med begrepet "reproduksjon".
Det er preget generasjonstid(tidsintervallet hvor antall celler dobles) og et slikt konsept som bakteriekonsentrasjon(antall celler i 1 ml).
I motsetning til den mitotiske delingssyklusen i eukaryoter, skjer reproduksjonen av de fleste prokaryoter (bakterier) ved binær fisjon, og actinomycetes - spirende. Imidlertid eksisterer alle prokaryoter i haploid tilstand, siden DNA-molekylet er representert i cellen i entall.
2. Når du studerer prosessen med bakteriell reproduksjon, er det ekstremt viktig å vurdere det bakterier eksisterer alltid i form av mer eller mindre tallrike populasjoner, og utvikling bakteriepopulasjon i et flytende næringsmedium i batchkultur kan betraktes som lukket system.
Det er 4 faser i denne prosessen:
‣‣‣ 1. - første, eller etterslep fase eller forsinket reproduksjonsfase - karakterisert begynnelsen av intensiv cellevekst, Men frekvensen av deres deling forblir lav;
‣‣‣ 2. - logaritmisk eller loggfase, eller eksponentiell fase, - karakterisert en konstant maksimal hastighet av celledeling og en betydelig økning i antall celler i befolkningen;
‣‣‣ 3. - stasjoner fase - kommer når antall celler i befolkningen slutter å øke. Dette skyldes det faktum at det oppstår en likevekt mellom antall nydannede og døende celler. Antall levende bakterieceller i en populasjon per volumenhet næringsmedium i stasjonær fase er betegnet som M-konsentrasjon. Denne indikatoren er et karakteristisk trekk for hver type bakterier;
‣‣‣ 4. - dø-off fase (logaritmisk død) - karakterisert en overvekt i populasjonen av antall døde celler og en progressiv nedgang i antall levedyktige celler i populasjonen. Opphør av vekst i antall (reproduksjon) av en populasjon av mikroorganismer skjer på grunn av uttømming av næringsmediet og/eller akkumulering av metabolske produkter fra mikrobielle celler i det. Av denne grunn, ved å fjerne metabolske produkter og/eller erstatte næringsmediet, regulere overgangen til den mikrobielle populasjonen fra den stasjonære fasen til den døende fasen, er det mulig å skape åpent biologisk system søker å eliminere dynamisk likevekt på et visst nivå av befolkningsutvikling.
Denne prosessen med å dyrke mikroorganismer kalles ofte flytdyrking (kontinuerlig kultur). Vekst i en kontinuerlig kultur gjør det mulig å oppnå store masser av bakterier under strømningsdyrking i spesielle apparater (kjemostater og turbidistater) og brukes i produksjon av vaksiner, samt i bioteknologi for produksjon av ulike biologisk aktive stoffer produsert av mikroorganismer .
For å studere metabolske prosesser gjennom celledelingssyklusen er det også mulig å bruke synkrone kulturer - slike bakteriekulturer, som alle medlemmer av befolkningen er i en fase av syklusen. Dette oppnås ved hjelp av spesielle dyrkingsmetoder.
I dette tilfellet, etter flere samtidige delinger, går den synkroniserte cellesuspensjonen gradvis tilbake til asynkron deling, slik at antallet celler deretter øker ikke trinnvis, men kontinuerlig.
3. Når det dyrkes på faste næringsmedier, dannes det bakterier kolonier - en klynge av bakterier av samme art synlig for det blotte øye, som oftest er avkom av én celle.
Kolonier av bakterier av forskjellige arter er forskjellige:
‣‣‣ form;
‣‣‣ størrelse;
‣‣‣ åpenhet;
‣‣‣ farge;
‣‣‣ høyde;
‣‣‣ overflatens og kantenes natur;
‣‣‣ konsistens.
Kolonienes natur - en av de taksonomiske egenskapene til bakterier.
44. Definisjon og essens av begrepene «biosfære» og «biocenose». Moderne ideer om utviklingen av mikrober.
I naturen bor mikroorganismer i nesten alle miljøer (jord, vann, luft) og er mye mer utbredt enn andre levende vesener. På grunn av mangfoldet av mekanismer for resirkulering av mat og energikilder, samt uttalt tilpasning til ytre påvirkninger, kan mikroorganismer leve der andre former for liv ikke kan overleve.
De naturlige habitatene til de fleste organismer er vann, jord og luft. Antallet mikroorganismer som lever på planter og i dyrekropper er mye mindre. Den brede distribusjonen av mikroorganismer er assosiert med den enkle spredningen gjennom luft og vann; spesielt er overflaten og bunnen av ferskvanns- og saltvannsforekomster, samt flere centimeter av det øverste jordlaget, fulle av mikroorganismer som ødelegger organisk materiale. Et mindre antall mikroorganismer koloniserer overflaten og noen indre hulrom hos dyr (for eksempel mage-tarmkanalen, øvre luftveier) og planter.
I habitatområder dannes det mikroorganismer biocenoser[fra gresk bios, liv, + koinos, fellesskap] - komplekse assosiasjoner med spesifikke og ofte uvanlige relasjoner. Hvert mikrobielt samfunn i en bestemt biocenose danner spesifikke autoktone mikroorganismer[fra gresk biler din, + chthon, land, lokalitet], det vil si mikrober som er iboende i et spesifikt område.
Symbiose[fra gresk symbiose, samliv] - den langsiktige sameksistensen av mikroorganismer i langlivede samfunn. Forholdet der en mikroorganisme befinner seg utenfor cellene til verten (større organisme) er kjent som ektosymbiose: når den er lokalisert inne i celler, er den kjent som endosymbiose. Typiske ektosymbiotiske mikrober - Escherichia coli slekter av bakterier Bacteroides Og Bifidobacterium, Proteus vulgaris, så vel som andre representanter for intestinal mikroflora. Som et eksempel på endosymbiose kan vi vurdere plasmider som gir for eksempel bakteriell resistens mot legemidler. Symbiotiske relasjoner er også delt inn i henhold til fordelene mottatt av hver partner.
Gjensidighet[fra lat. mutuus, gjensidig] - gjensidig fordelaktig symbiotisk forhold. Således produserer mikroorganismer biologisk aktive stoffer som er nødvendige for vertskroppen (for eksempel B-vitaminer). Samtidig er endo- og ektosymbionter som lever i makroorganismer beskyttet mot ugunstige miljøforhold (tørking og ekstreme temperaturer) og har konstant tilgang på næringsstoffer. Av alle typer gjensidighet er den mest overraskende dyrkingen av noen sopp av insekter (biller og termitter). På den ene siden bidrar dette til en bredere spredning av sopp, på den andre - o\
mj gir en konstant kilde til næringsstoffer for larvene. Dette minner om menneskelig dyrking av nyttige planter og mikroorganismer.
Kommensalisme- en type symbiose der bare én partner drar nytte av (uten å forårsake "synlig" skade på den andre); mikroorganismer involvert i slike forhold er kommensaler [fra lat. celle-, s, + mensa, bord; bokstavelig talt - bordkamerater]. Kommensale mikroorganismer koloniserer huden og hulrommene i menneskekroppen (for eksempel mage-tarmkanalen), uten å forårsake "synlig" skade; deres helhet er normal mikrobiell flora (naturlig mikroflora). Typiske ektosymbiotiske kommensale organismer er Escherichia coli, bifidobakterier, stafylokokker og laktobaciller. Mange kommensale bakterier tilhører opportunistisk mikroflora og er i stand til, under visse omstendigheter, å forårsake sykdommer i makroorganismen (for eksempel når de introduseres i blodet under medisinske prosedyrer).
Vekst og reproduksjon av bakterier. Mekanisme og reproduksjonshastighet. Faser av mikrobiell reproduksjon. - konsept og typer. Klassifisering og funksjoner i kategorien "Vekst og reproduksjon av bakterier. Mekanisme og reproduksjonshastighet. Faser av mikrobiell reproduksjon." 2017, 2018.
Bakterievekst - Dette er en økning i bakteriecellestørrelse uten å øke antall individer i befolkningen. Cellevekst er ikke ubegrenset. Etter å ha nådd en kritisk størrelse, gjennomgår cellen deling.
Reproduksjon av bakterier - en prosess som sikrer en økning i antall individer i en populasjon. Bakterier er preget av høy reproduksjonshastighet.
Vekst går alltid foran reproduksjon. Bakterier formerer seg ved tverrgående binær fisjon, der to identiske datterceller dannes fra en morcelle. Hos de fleste gram-positive bakterier skjer deling gjennom syntesen av en tverrgående skillevegg som går fra periferien til sentrum. Cellene til de fleste gramnegative bakterier deler seg ved innsnevring.
Prosessen med bakteriell celledeling begynner med replikering av kromosomalt DNA. Replikering begynner i en valgt region, kalt opprinnelse, som har en spesifikk nukleotidsekvens. En eller to replikasjonsgafler kan forekomme her. Mer enn 20 enzymer er involvert i replikasjonsprosessen. Siden bakterielt DNA er dobbelttrådet, må det deles før replikasjon. Denne prosessen involverer enzymene helicase, som vikler ut dobbelthelixen, og topoisomerase, som forhindrer dannelsen av sekundære krøller. SSB-protein binder seg til enkelttrådet DNA, og hindrer det i å refolde seg til en dobbel helix. Som et resultat dannes en replikasjonsgaffel. Syntesen av nye DNA-kjeder utføres av enzymet DNA polymyrase. For å utføre polymerisasjonsreaksjonen av nukleotider på malen til moderkjeden, krever polymerase en primer. Primeren er en kort RNA-nukleotidkjede, komplementær til malstrengen, med en fri 3/- ende. Etter at DNA-strengen har begynt å syntetiseres, fjernes RNA-primeren. Siden DNA-trådene i en dupleks er antiparallelle, faller retningen for avvikling av dobbeltstrengen bare sammen med retningen for DNA-syntese på en mal, som kalles den ledende. På den komplementære strengen syntetiseres DNA i korte fragmenter, som deretter sys sammen til en streng av DNA-ligaser. Den bakterielle DNA-replikasjonsprosessen fortsetter til alt DNA er doblet.
Når bakterier tilsettes næringsmediet, vokser og formerer de seg til innholdet av en av de nødvendige komponentene i mediet når et minimum, hvoretter vekst og reproduksjon stopper. Hvis vi ikke tilfører næringsstoffer og ikke fjerner sluttprodukter av stoffskiftet, får vi en statistisk bakteriekultur.
Bakterielle reproduksjonsfaser:
1. Første(lagfase) dekker tidsperioden fra det øyeblikket bakteriene inokuleres til starten av veksten. Dens varighet er i gjennomsnitt 2-5 timer og avhenger av sammensetningen av næringsmediet.
2. Eksponentiell(logaritmisk) fase. Karakterisert av en konstant maksimal hastighet av celledeling. Denne hastigheten avhenger av bakterietypen og næringsmediet. Tiden det tar før cellene dobles kalles generasjonstid. Denne tiden varierer fra flere minutter til flere timer.
3. Stasjonær fase oppstår når antall celler slutter å øke. Med en reduksjon i konsentrasjonen av næringsstoffer i næringsmediet, en reduksjon i partialtrykket av oksygen og akkumulering av giftige metabolske produkter, reduseres veksthastigheten til bakterier. Varigheten av den stasjonære fasen er flere timer og avhenger av bakterietypen.
4. Nedgangsfase oppstår på grunn av akkumulering av sure metabolske produkter eller som et resultat av autolyse under påvirkning av egne enzymer. Varigheten av denne fasen varierer fra ti timer til flere uker.
3.2. Næringsmedier, prinsipper for deres klassifisering, krav til næringsmedier, betingelser for dyrking av mikroorganismer.
Grunnlaget for bakteriologisk arbeid er næringsmedier, som ofte bestemmer resultatene av studien etter deres kvalitet.
Grunnleggende krav til næringsmedier:
1. Kulturmedier må inneholde alle de næringsstoffene som er nødvendige for å mate mikroben, dvs. har næringsverdi.
2. Ha tilstrekkelig fuktighet
3. Ha en optimal pH (7,2-7,6) surhet i miljøet.
4. Vær isotonisk (NaCl-konsentrasjon 0,87%), for halofile bakterier er saltkonsentrasjonen 1% eller høyere.
5. Ha et optimalt elektronisk potensial, som indikerer innholdet av oppløst oksygen i mediet. Den skal være høy for aerobe og lav for anaerobe.
6. Vær gjennomsiktig slik at bakterievekst er synlig, spesielt i flytende medier.
7. Vær steril (slik at det ikke er andre bakterier).
For å tilberede næringsmedier brukes produkter av animalsk opprinnelse (kjøtt, fisk, blod, egg, melk) og produkter av vegetabilsk opprinnelse (poteter). Syntetiske næringsmedier sammensatt av kjemiske forbindelser brukes også.
Kilden til nitrogen for bakterier er enkle ammoniumforbindelser, aminosyrer eller peptoner; karbonkilde – sukker, flerverdige alkoholer, organiske syrer. Bakteriens behov for uorganiske grunnstoffer dekkes av salter tilsatt næringsmediet: NaCl, KN 2 PO 4, K 2 HPO 4.
Avhengig av konsistensen kan næringsmedier være: flytende, halvflytende og tett. Mediets tetthet oppnås ved å tilsette agar. Agar er et polysakkarid hentet fra alger. Den smelter ved en temperatur på 100 oC, avkjøles ved en temperatur på 45-50 oC. For halvflytende medier tilsettes agar i en konsentrasjon på 0,5%, for tette medier - 1,5-2%. Flytende medier inneholder ikke agar-agar.
Sammensetningen av næringsmedier kan være enkelt og komplekst. Enkle medier inkluderer peptonvann, kjøttpeptonbuljong, kjøttpeptonagar, Hottinger-agar. Komplekse er enkle med tillegg av en ekstra ernæringskomponent (sukker, myse, gallebuljonger, blod, myse, eggeplomme-salt-agarer, Keith-Tarozzi, Wilson-Blair-medium).
Avhengig av formålet med miljøet er de delt inn i:
1. Generelt formål – for dyrking av de fleste bakterier (kjøttpeptonagar, blodagar).
2. Spesielt formål:
a) valgfrie miljøer– dette er mediet som en spesifikk mikroorganisme vokser på. For eksempel brukes alkalisk agar med en pH på 9 for å isolere Vibrio cholerae.
b) Med anrikningstrinn– dette er miljøer som stimulerer veksten av en bestemt mikroorganisme, og hemmer veksten til andre. For eksempel stimulerer magnesium- og selenittmedier veksten av bakterier av slekten Salmonella, og hemmer veksten av E. coli.
c) differensialdiagnostiske miljøer tjene til å studere den enzymatiske aktiviteten til bakterier (Hiss-medium).
d) kombinerte næringsmedier kombinere et selektivt medium som undertrykker veksten av medfølgende flora og et differensialdiagnostisk medium (Ploskirevs medium for isolering av Shigella, vismut-sulfittagar for Salmonella). Begge disse mediene hemmer veksten av E. coli.
For å skille prototrofe og auxotrofe bakterier brukes selektive medier. Prototrofer vokser på et minimalt medium som bare inneholder salter og karbohydrater, siden de selv er i stand til å syntetisere metabolittene de trenger for utvikling. Auxotrofer trenger medier som inneholder visse aminosyrer, vitaminer, dvs. vekstfaktorer.
Tilberedning av næringsmedier er et av de viktigste og vanskeligste områdene innen bakteriologisk arbeid.
For tiden har den medisinske industrien organisert produksjonen av hermetiserte medier. Tørre næringsmedier er inneholdt i plastbeholdere med tettsittende lokk for å sikre en lufttett forsegling.
Bakteriedyrkingsforhold:
1. Tilgjengelighet av et komplett næringsmedium.
2. En viss dyrkingstemperatur (optimal temperatur 37 0 C).
3. En viss kultiveringsatmosfære. Strenge aerober krever oksygen, så de vokser godt på overflaten av agar-petriskåler eller i et tynt topplag av flytende medium. For vekst av aerober i det dype laget av et flytende medium, er det nødvendig å kontinuerlig blande eller riste næringsmediene slik at oksygen fordeles gjennom hele volumet av mediet. For fakultative anaerober brukes de samme metodene. Mikroaerofile formerer seg ved redusert partialtrykk av oksygen. CO 2 -konsentrasjonen bør være 1-5 %. For å gjøre dette brukes spesielle CO 2 inkubatorer eller avlingene plasseres i ekssikkatorer der et varmt stearinlys er installert. For vekst av obligatoriske anaerober er det nødvendig å utelukke tilgang til oksygen. For å gjøre dette, tilsett oksygenreduserende stoffer (tioglykolsyre) til næringsmediene, regenerer flytende næringsmedier fra luftoksygen ved å koke dem, bruk oksygenabsorbere ved å plassere dem i hermetisk forseglede "gaspack"-beholdere og bruk anaerostater.
4. Dyrkingstid (18-48 timer). For dyrking av Mycobacterium tuberculosis (3-4 uker).
5. Belysning. Lys er nødvendig for å dyrke fototrofe bakterier.
Under industrielle forhold, for å oppnå biomasse av bakterier eller sopp for å få antibiotika, vaksiner og diagnostiske medisiner, utføres dyrking i apparater (fermentorer) med streng overholdelse av de optimale parameterne for vekst og forplantning av avlinger.
Ernæring av bakterier.
Ernæring refererer til prosessene for inn- og utgang av næringsstoffer inn og ut av celler. Ernæring sørger først og fremst for celleproduksjon og metabolisme.
Essensielle næringsstoffer inkluderer: karbon, oksygen, hydrogen, nitrogen, fosfor, kalium, magnesium, kalsium. I tillegg til organogener er mikroelementer nødvendige. De gir enzymaktivitet. Disse er sink, mangan, molybden, kobolt, kobber, nikkel, wolfram, natrium, klor.
Bakterier har en rekke kilder til næringsstoffer.
Avhengig av kilden til karbon, er bakterier delt inn i: 1) autotrofer (bruk uorganiske stoffer - CO 2); 2) heterotrofer (bruk organiske C-heksoser, flerverdige alkoholer, aminosyrer);
Ernæringsprosesser må sørge for energibehovet til bakteriecellen. Basert på energikilder deles mikroorganismer inn i: 1) fototrofer - kilden er solenergi; 2) kjemotrofer - få energi gjennom redoksreaksjoner; 3) kjemolitotrofer - bruk uorganiske forbindelser; 4) kjemoorganotrofer - bruk organiske stoffer.
Medisinsk mikrobiologi studerer bakterier som er heterokjemoorganotrofer.
Bakterielle vekstfaktorer er vitaminer, aminosyrer, purin- og pyrimidinbaser, hvis tilstedeværelse akselererer veksten. Blant bakterier er det: 1) prototrofer (i stand til å syntetisere de nødvendige stoffene selv); 2) auxotrofer (trenger vekstfaktorer).
Mikroorganismer assimilerer næringsstoffer i form av små molekyler, så proteiner, polysakkarider og andre biopolymerer kan tjene som ernæringskilder først etter at de er brutt ned av eksoenzymer til enklere forbindelser.
Veier for inntreden av metabolitter og ioner i den mikrobielle cellen: I. Passiv transport (uten energikostnader): enkel diffusjon; 2) forenklet diffusjon (langs en konsentrasjonsgradient, ved hjelp av bærerproteiner). II. Aktiv transport (med energiforbruk, mot en konsentrasjonsgradient; i dette tilfellet samhandler substratet med et bærerprotein på overflaten av den cytoplasmatiske membranen).
Det er modifiserte versjoner av aktiv transport - overføring av kjemiske grupper. Fosforylerte enzymer fungerer som bærerproteiner, så substratet transporteres i en fosforylert form. Denne overføringen av en kjemisk gruppe kalles translokasjon.
Begreper om bakterievekst og reproduksjon
For mikrobiologisk diagnostikk, studiet av mikroorganismer og for bioteknologiske formål dyrkes mikroorganismer på kunstige næringsmedier.
Under bakterievekst forstå en økning i cellemasse uten å endre antallet i en populasjon som et resultat av koordinert reproduksjon av alle cellulære komponenter og strukturer. En økning i antall celler i en populasjon av mikroorganismer er betegnet med begrepet "reproduksjon". Det er preget av generasjonstid (tidsintervallet hvor antall celler dobles) og et konsept som bakteriekonsentrasjon (antall celler i 1 ml).
I motsetning til den mitotiske delingssyklusen hos eukaryoter, skjer reproduksjonen av de fleste prokaryoter (bakterier) ved binær fisjon, og av aktinomyceter ved knoppskyting. Dessuten eksisterer alle prokaryoter i en haploid tilstand, siden DNA-molekylet er representert i cellen i entall.
Bakteriepopulasjon
Når man studerer prosessen med bakteriell reproduksjon, er det nødvendig å ta hensyn til at bakterier alltid eksisterer i form av mer eller mindre tallrike populasjoner, og utviklingen bakteriepopulasjon i et flytende næringsmedium i batchkultur kan betraktes som et lukket system. Det er 4 faser i denne prosessen:
- 1. - første, eller etterslep fase eller forsinket reproduksjonsfase- preget av begynnelsen av intensiv cellevekst, men hastigheten på deres deling forblir lav;
- 2. - logaritmisk eller loggfase, eller eksponentiell fase,- preget av en konstant maksimal hastighet av celledeling og en betydelig økning i antall celler i befolkningen;
- 3. - stasjoner fase- oppstår når antall celler i en populasjon slutter å øke. Dette skyldes det faktum at det oppstår en likevekt mellom antall nydannede og døende celler. Antall levende bakterieceller i en populasjon per volumenhet næringsmedium i stasjonær fase betegnes som M-konsentrasjon. Denne indikatoren er et karakteristisk trekk for hver type bakterier;
- 4. - dø-off fase (logaritmisk død)- preget av en overvekt i befolkningen av antall døde celler og en progressiv nedgang i antall levedyktige celler i befolkningen. Opphør av vekst i antall (reproduksjon) av en populasjon av mikroorganismer skjer på grunn av uttømming av næringsmediet og/eller akkumulering av metabolske produkter fra mikrobielle celler i det. Derfor, ved å fjerne metabolske produkter og/eller erstatte næringsmediet, regulere overgangen til den mikrobielle populasjonen fra den stasjonære fasen til den døende fasen, er det mulig å skape et åpent biologisk system som har en tendens til å eliminere dynamisk likevekt på et visst nivå av befolkningsutvikling.
Denne prosessen med å dyrke mikroorganismer kalles flytdyrking (kontinuerlig kultur). Vekst i en kontinuerlig kultur gjør det mulig å oppnå store masser av bakterier under strømningsdyrking i spesielle apparater (kjemostater og turbidistater) og brukes i produksjon av vaksiner, samt i bioteknologi for produksjon av ulike biologisk aktive stoffer produsert av mikroorganismer .
For å studere metabolske prosesser gjennom celledelingssyklusen er det også mulig å bruke synkrone kulturer- slike bakteriekulturer, hvor alle medlemmer av befolkningen er i samme fase av syklusen. Dette oppnås ved hjelp av spesielle dyrkingsmetoder.
Men etter flere samtidige delinger går den synkroniserte cellesuspensjonen gradvis tilbake til asynkron deling, slik at antall celler ikke lenger øker trinnvis, men kontinuerlig.
Kolonier
Når det dyrkes på faste næringsmedier, dannes bakterier kolonier- en klynge av bakterier av samme art synlig for det blotte øye, som oftest er avkom fra én celle.
Kolonier av bakterier av forskjellige arter er forskjellige:
- form;
- størrelse;
- åpenhet;
- farge;
- høyde;
- naturen til overflaten og kantene;
- konsistens.
Kolonienes natur er en av de taksonomiske egenskapene til bakterier.
De intense prosessene med anabolisme og katabolisme i cellen fører til rask cellevekst.
Bakteriell vekst er en ordnet økning i antall og størrelse på alle komponentene i cellen, med forbehold om tilstedeværelsen av alle nødvendige kjemiske elementer, noe som fører til en økning i massen. Næringssubstrater må inneholde disse elementene i en metabolsk tilgjengelig form. Cellevekst er ikke ubegrenset. Etter å ha nådd en kritisk størrelse, gjennomgår cellen deling eller reproduksjon.
De fleste bakterier deler seg ved tverrgående binær fisjon eller cytokinese. Hos de fleste gram-positive bakterier skjer deling gjennom syntesen av en tverrgående skillevegg som går fra periferien til sentrum. Cellene til de fleste gramnegative bakterier deler seg ved innsnevring. Delingsprosessen gjentas med omtrent like intervaller (fra flere minutter til flere dager), som er en individuell genetisk karakteristikk av den mikrobielle arten. Som et resultat av reproduksjon øker antallet celler i befolkningen kraftig.
Reproduksjon eller reproduksjon i bakterier er delingen av supercoiled nukleoid-DNA i to dattertråder, som hver fullføres ytterligere av en komplementær tråd og dannelsen av to datterceller skjer samtidig (semi-konservativ metode).
Reproduksjon er preget generasjonstid(tidsintervallet hvor antall celler dobles) og et slikt konsept som bakteriekonsentrasjon(antall celler i 1 ml).
Når bakterier introduseres i et næringsmedium, vokser de og formerer seg til innholdet av noen av de nødvendige komponentene i mediet når et minimum, hvoretter vekst og reproduksjon stopper. Hvis vi gjennom hele denne tiden ikke tilfører næringsstoffer og ikke fjerner sluttproduktene av stoffskiftet, får vi statisk bakteriekultur. En statisk (batch) kultur av bakterier oppfører seg som en flercellet organisme, med genetisk vekstbegrensning. Konstruerer vi en graf med tid på abscisseaksen og antall celler på ordinataksen, får vi en kurve som beskriver avhengigheten av antall dannede celler på reproduksjonstidspunktet, som kalles vekstkurve.
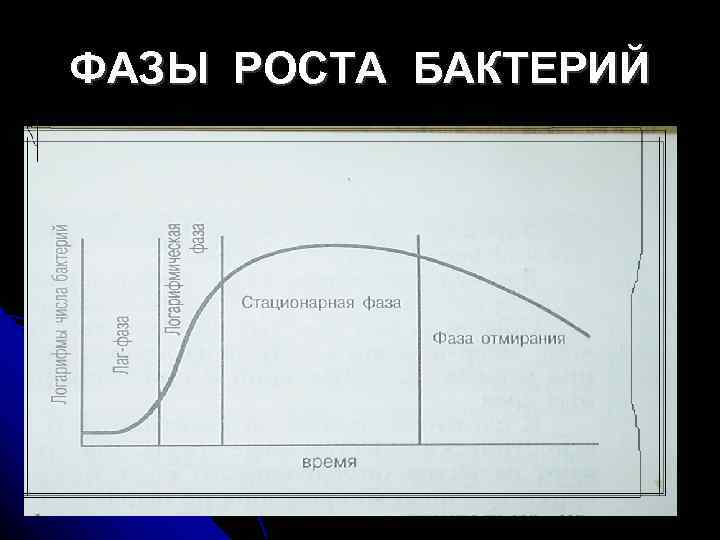
Vekstkurve for bakterier i et næringsmedium. På denne kurven kan flere faser skilles ut, som erstatter hverandre i en bestemt rekkefølge (fig. 11):
1. Innledende - lagfase(Engelsk) etterslep- å henge etter). Dekker tidsrommet mellom inokulering av bakterier og start av reproduksjon. Dens varighet er i gjennomsnitt 2-5 timer og avhenger av sammensetningen av næringsmediet og alderen på avlingen som sås. Under lagfasen tilpasser bakterieceller seg til nye dyrkingsforhold og induserbare enzymer syntetiseres.
2. Eksponentiell (logaritmisk) fase. Karakterisert av en konstant maksimal hastighet av celledeling, en fase med geometrisk vekst med en kraftig økning i populasjonen av mikroorganismer (2 i grad n). Reproduksjonshastigheten avhenger av bakterietypen og næringsmediet. Celledoblingstiden kalles generasjonstid, som varierer avhengig av type bakteriekultur: i bakterier av slekten Pseudomonas det er lik 14 minutter, og Mycobacterium 18 - 24 timer Størrelsen på cellene og proteininnholdet i dem forblir konstant under eksponentiell fase. Bakteriekulturen i denne fasen består av standardceller.
Ris. 11. Faser av bakteriell reproduksjon
3. Stasjonær fase(fase av likevekt av reproduksjon og død av mikrobielle celler). Oppstår når antall celler slutter å øke. Siden veksthastigheten avhenger av konsentrasjonen av næringsstoffer, når innholdet av næringsstoffer i næringsmediet synker, avtar også veksthastigheten. En reduksjon i veksthastighet oppstår også på grunn av den høye tettheten av bakterieceller, en reduksjon i partialtrykket av oksygen og akkumulering av giftige metabolske produkter. Varigheten av den stasjonære fasen er flere timer og avhenger av typen bakterier og egenskapene til dyrkingen deres.
4. Nedgangsfase eller død - en reduksjon i populasjonsstørrelse på grunn av en reduksjon og mangel på forhold for reproduksjon av mikroorganismer. Oppstår på grunn av akkumulering av sure metabolske produkter eller som et resultat av autolyse under påvirkning av egne enzymer. Varigheten av denne fasen varierer fra ti timer til flere uker.
Denne dynamikken er typisk for periodiske avlinger med gradvis uttømming av næringsstoffer og akkumulering av metabolitter. Den konstante tilstedeværelsen av bakteriepopulasjonen i den logaritmiske vekstfasen observeres i en kontinuerlig kultur, som oppnås ved gradvis dosering av næringsstoffer, kontroll av tettheten til bakteriesuspensjonen og fjerning av metabolitter. Denne prosessen med å dyrke mikroorganismer kalles flytdyrking (kontinuerlig kultur). Vekst i en kontinuerlig kultur gjør det mulig å oppnå store masser av bakterier under strømningsdyrking i spesielle apparater (kjemostater og turbidistater) og brukes i produksjon av vaksiner, samt i bioteknologi for produksjon av ulike biologisk aktive stoffer produsert av mikroorganismer .
 Russiske studenter vant ICPC World Programming Championship
Russiske studenter vant ICPC World Programming Championship "Denisovan-mannen" "arvet" mest av alt i genomet til asiater
"Denisovan-mannen" "arvet" mest av alt i genomet til asiater Handling for å skremme sjekkpunkter i Chelyabinsk Hva er konseptet offentlig sikkerhet
Handling for å skremme sjekkpunkter i Chelyabinsk Hva er konseptet offentlig sikkerhet