Көз торындағы фотохимиялық реакциялар. Тор қабықшасының рецепторларындағы фотохимиялық реакциялар Фотохимиялық реакцияның кванттық шығымы
Көз торына жарық әсер еткенде, химиялық өзгерістертаяқшалар мен конустардың сыртқы сегменттерінде кездесетін пигменттер. Болғандықтан фотохимиялық реакцияфоторецепторлардың қозуы жүреді көз торы.
Фотосезімтал пигменттер жануарлардың көзінің торлы қабығында өткен ғасырдың 70-жылдарының соңында табылған және бұл заттардың жарықта өшетіндігі көрсетілген. Адамдардың және көптеген жануарлардың торлы таяқшаларында родопсин пигменті немесе көрнекі күлгін бар, оның құрамы, қасиеттері және химиялық өзгерістері соңғы онжылдықтарда егжей-тегжейлі зерттелген (Wold et al.). Йодопсин пигменті құстардың конустарында табылған. Шамасы, конустарда жарыққа сезімтал басқа пигменттер де бар. Руштон конустарда пигменттердің болуын көрсетеді - хлоролаб және эритролаб; олардың біріншісі жасылға, ал екіншісі - спектрдің қызыл бөлігіне сәйкес сәулелерді жұтады.
Родопсинретиннен, А витаминінің альдегидінен және опсин протеинінен тұратын жоғары молекулалы қосылыс болып табылады. Жарық әсер еткенде бұл заттың химиялық түрлену циклі жүреді. Жарықты сіңіре отырып, ретинен өзінің геометриялық изомеріне айналады, оның бүйірлік тізбегінің түзетілуімен сипатталады, бұл ретинен мен ақуыз арасындағы байланыстың бұзылуына әкеледі. Бұл жағдайда алдымен кейбір аралық заттар – лумпродопсин және метаходопсин түзіледі, одан кейін опсиннен ретинен бөлінеді. Ретиненредуктаза деп аталатын ферменттің әсерінен соңғысы таяқшалардың сыртқы сегменттерінен пигмент қабатының жасушаларына түсетін А витаминіне айналады.
Көздер қараңғыланған кезде көрнекі күлгін регенерация жүреді, яғни родопсиннің ресинтезі. Бұл процесс көз торының А витаминінің цис изомерін алуын талап етеді, одан ретинен түзіледі. Ағзада А дәрумені болмаған жағдайда родопсиннің түзілуі күрт бұзылады, бұл жоғарыда аталған түнгі соқырлықтың дамуына әкеледі. А витаминінен ретиненнің түзілуі ферменттік жүйенің қатысуымен жүретін тотығу процесі болып табылады. Тотығу процестері бұзылған сүтқоректілердің оқшауланған тор қабығында родопсиннің қалпына келуі болмайды.
Көз торындағы фотохимиялық процестерөте үнемді түрде пайда болады, яғни өте жарқын жарықтың әсерінен таяқшаларда болатын родопсиннің аз ғана бөлігі ғана ыдырайды. Осылайша, Вальдтың айтуынша, 100 люкс интенсивтілігі бар жарық әсер еткенде, 5 секундтан кейін ондағы осы заттың 18 миллион молекуласының әрбір таяқшасында визуалды күлгіннің тек 1200 молекуласы ыдырайды, яғни шамамен 0,005%. родопсин ыдырайды.
Родопсиннің жарықты жұтуы және оның бөлінуі оған әсер ететін жарық сәулелерінің толқын ұзындығына байланысты әр түрлі болады. Адамның тор қабығынан алынған родопсин спектрдің жасыл бөлігінде жатқан толқын ұзындығы шамамен 500 ммК жарық сәулелерінің әсерінен максималды сіңіруді көрсетеді. Дәл осы сәулелер қараңғыда ең жарқын болып көрінеді. Қараңғыда жарықтың жарықтығын субъективті бағалау қисығымен әртүрлі толқын ұзындықтағы жарық әсерінен родопсиннің жұтылу және ағарту қисығын салыстыру олардың толық сәйкестігін көрсетеді ( күріш. 215).
|
Егер сіз торлы қабықты алюминий ерітіндісімен өңдесеңіз, яғни оны бекітетін болсаңыз, онда бұл родопсинді одан әрі ыдыраудан қорғайды және торда сіз бұрын көз қараған объектінің бейнесін көре аласыз (оптограмма деп аталады). Йодопсиннің құрылысы родопсинге жақын. Йодопсин сонымен қатар конустарда түзілетін және таяқша опсиннен ерекшеленетін опсин белогымен ретиненнің қосылысы болып табылады. Родопсин мен йодопсиннің жарықты жұтуы әртүрлі. Йодопсин спектрдің сары сағатында жататын толқын ұзындығы шамамен 560 мм болатын жарық сәулелерін барынша сіңіреді. Күріш. 215. Қараңғыда адам көзінің сезімталдығын көрнекі күлгіннің жұтылу спектрімен салыстыру. Нүктелер сезімталдықты көрсетеді. |
Студент міндетті
білу:
1. Жүйке жүйесінің электрлік импульстері. Рефлекторлық доға.
2. Бұлшықеттің жиырылу механизмі. Ас қорыту.
3. Оттегінің берілуі және сіңірілуі. Қан мен лимфаны тазарту.
білуұғымдарға анықтама беріңіз:импульс, бұлшықет, қан, лимфа.
Ағзадағы дәнекер ұлпасының түрлері. Дәнекер ұлпаның қызметтері. Сүйек тіні. Шеміршек ұлпасы. Қан және лимфа. Майлы тін. Май ұлпасының қызметтері. Бұлшық ет ұлпасы және оның түрлері. Тегіс бұлшықет тіні. Жолақты бұлшықет тіні. Жүрек (жүрек бұлшықеті). Бұлшық ет ұлпасының қызметтері. Жүйке ұлпасы. Жүйке жасушалары (нейрондар) және жасушааралық зат – нейроглия. Жүйке ұлпасының қызметтері.
Тақырып 36. Тірі организмдегі (адам ағзасындағы) электромагниттік құбылыстар: жүрек пен мидың электрлік ырғақтары, жүйке импульстарының электрлік табиғаты.
Студент міндетті
білу:
1. Тұжырымдама электромагниттік құбылыстірі организмде.
2. Ырғақ туралы түсінік. Мидың электрлік ырғақтары.
3. Фибрилляция және дефибрилляция.
білуұғымдарға анықтама беріңіз:
Тақырып 37. Көру құбылысы: оптика, фотохимиялық реакциялар, ақпаратты талдау.
Студент міндетті
білу:
1. Көру туралы түсінік.
2. Ми және көру.
білуұғымдарға анықтама беріңіз:көру, нервтер, линзалар, көз торы.
Көздегі фотохимиялық реакциялар. Ақпаратты талдау механизмі.
Тақырып 38. Электромагниттік толқындар мен радиоактивті сәулелердің адам ағзасына әсері.
Студент міндетті
білу:
1. Адам денесінің электромагниттік өрісі (ЭМӨ).
2. Жердің ЭҚК-нің биологиялық әсері, технологиясы.
3. Электромагниттік түтін және оның әсері.
білуұғымдарға анықтама беріңіз:ЭҚК, радиоактивті сәулелену.
Мазмұны оқу материалы(дидактикалық бірлік):Адам денсаулығы үшін қауіпсіз электромагниттік өріс қарқындылығының шегі - 0,2 мкТ (microTesla). Тұрмыстық құрылғылар мен көліктерден электромагниттік өрістердің қарқындылығы. Радиоактивті сәулелену: альфа, бета, гамма сәулелену. Олардың адамға әсер ету механизмі. Адамдарды электромагниттік толқындар мен радиоактивті сәулелердің зиянды әсерінен қорғау әдістері мен құралдары.
Тақырып 39. Адам ағзасындағы макромолекулалардың рөлі, ферменттер және ферментативті реакциялар.
Студент міндетті
білу:
1. Адам ағзасындағы макромолекулалардың түрлері. Олардың физиологиялық процестерге әсері.
2. Фермент туралы түсінік.
3. Ферментативті реакциялар.
білуұғымдарға анықтама беріңіз:макромолекула, фермент.
Тақырып 40. Тұқым қуалайтын заңдылықтар. Адам геномы.
Студент міндетті
білу:
1. Хромосомалар мен ДНҚ-ның ашылуы.
2. Тұқым қуалайтын заңдылықтар.
3. Ғылыми-техникалық прогресс және адамзаттың генотипі.
білуұғымдарға анықтама беріңіз:ДНҚ, хромосома, генотоп.
Тақырып 41. Генетикалық анықталған аурулар және оларды емдеу мүмкіндіктері.
Студент міндетті
білу:
1. Тұқым қуалайтын ауру туралы түсінік.
2. Генетикалық анықталған ауруларды емдеу әдістері.
білуұғымдарға анықтама беріңіз:ауру, мутация.
Люминесценция құбылысы бұрыннан белгілі - зат белгілі бір жиіліктегі жарықты жұтады және өзі басқа жиіліктегі шашыраңқы сәуле шығарады 19 ғасырда Стокс ережені бекітті - шашыраңқы жарық жиілігі жиіліктен аз. жұтылатын жарық (ν жұту > ν dis) құбылыс түскен жарықтың жеткілікті жоғары жиілігі кезінде ғана болады.
Кейбір жағдайларда люминесценция дерлік инерциясыз жүреді - ол бірден пайда болады және жарықтандыру тоқтатылғаннан кейін 10 -7 -10 -8 с кейін тоқтайды. Бұл люминесценцияның ерекше жағдайы кейде деп аталады флуоресценция.Бірақ бірқатар заттардың (фосфор және басқалары) ұзақ уақытқа созылатын (біртіндеп әлсіреу) минуттар және тіпті сағаттар бар. Люминесценцияның бұл түрі деп аталады фосфоресценция.Қыздыру кезінде дене фосфорлану қабілетін жоғалтады, бірақ люминесценция қабілетін сақтайды.
Стокс ережесін өрнектейтін теңсіздіктің екі жағын да Планк тұрақтысына көбейтіп, мынаны аламыз:
Демек, атом жұтқан фотонның энергиясы ол шығаратын фотонның энергиясынан үлкен болады; Сонымен, жарық жұту процестерінің фотондық табиғаты мұнда да көрінеді.
Стокс ережесінен бар ауытқуларды кейінірек қарастырамыз (§ 10.6).
Фотохимия құбылыстарында - химиялық реакцияларжарықтың әсерінен - реакцияның жүруіне қажетті ең төменгі жиіліктің болуын анықтауға болады. Бұл фотондық тұрғыдан өте түсінікті: реакция болуы үшін молекула жеткілікті қосымша энергия алуы керек. Көбінесе құбылыс қосымша әсерлермен жасырылады. Осылайша, H 2 сутегі мен хлор Cl 2 қоспасы қараңғыда ұзақ уақыт болатыны белгілі. Бірақ жеткілікті жоғары жиіліктегі жарықпен әлсіз жарықтандыру кезінде қоспасы өте тез жарылып кетеді.
Оның себебі қайталама реакциялардың пайда болуында. Фотонды жұтқан сутегі молекуласы диссоциациялануы мүмкін (негізгі реакция):
H 2 +hν -> H + H.
Атом сутегі молекулалық сутектен әлдеқайда белсенді болғандықтан, жылу бөлінуімен екінші реттік реакция жүреді:
H+Cl 2 =HCl+Cl.
Осылайша, H және Cl атомдары босатылады. Олар C1 2 және H 2 молекулаларымен әрекеттеседі және фотондардың аз санының жұтылуымен қоздырылған кезде реакция өте жылдам өседі.

Әртүрлі фотохимиялық реакциялардың ішінде фотосуретке түсіру процесінде болатын реакциялар назар аударуға тұрарлық. Камера фотохимиялық реакцияларға қабілетті күміс бромиді бар фотографиялық эмульсия қабатында нақты (әдетте қысқартылған) кескінді жасайды. Реакцияланған молекулалардың саны шамамен жарықтың қарқындылығына және оның әсер ету уақытына пропорционалды (суретке түсіру кезіндегі экспозиция уақыты). Дегенмен, бұл сан салыстырмалы түрде өте аз; пайда болған «жасырын бейне» сәйкес әсер еткенде даму процесіне ұшырайды химиялық реагенттеркүміс бромидінің қосымша бөлінуі фотохимиялық реакция кезінде түзілетін орталықтарда жүреді. Содан кейін кескінді бекіту (бекіту) процесі жүреді: реакцияға түспеген жарыққа сезімтал күміс бромиді ерітіндіге ауысады және фотоқабатта металл күміс қалады, бұл алынған теріс кескіннің жеке аймақтарының мөлдірлігін анықтайды (жарық неғұрлым көп сіңірілсе, сәйкес аймақ қараңғырақ). Фотоқағазды (немесе пленканы) негатив арқылы жарықтандыру арқылы қағазда (ол әзірленгеннен және бекітілгеннен кейін) фотосуретке түсірілетін объектіге сәйкес жарықтың таралуы алынады (әрине, егер фотосуретті түсіру және өңдеу үшін тиісті жағдайлар болса). материал кездеседі). Түсті фотосуретте фильм спектрдің үш түрлі бөлігіне сезімтал үш қабаттан тұрады.
Бұл қабаттар бір-біріне жарық сүзгілері ретінде қызмет етеді және олардың әрқайсысының жарықтандыруы спектрдің белгілі бір бөлігімен ғана анықталады. Ақ-қара фотографиялық процеске қарағанда әлдеқайда күрделі, түсті фотосурет процесі негізінен біріншіден еш айырмашылығы жоқ және әдеттегі фотоникалық процесс болып табылады.
Көру анализаторы – толқын ұзындығы 400-700 нм болатын электромагниттік сәулелену және фотондардың дискретті бөлшектері немесе кванттар түрінде жарық энергиясын қабылдайтын және көрнекі сезімдерді қалыптастыратын құрылымдардың жиынтығы. Көздің көмегімен бізді қоршаған әлем туралы барлық ақпараттың 80-90% қабылданады.
Күріш. 2.1
Көру анализаторының белсенділігінің арқасында олар заттардың жарықтандырылуын, олардың түсін, пішінін, өлшемін, қозғалыс бағытын және олардың көзден және бір-бірінен қашықтығын ажыратады. Мұның бәрі кеңістікті бағалауға, айналаңыздағы әлемді шарлауға және әртүрлі мақсатты әрекеттерді орындауға мүмкіндік береді.
Көру анализаторы ұғымымен қатар көру мүшесі деген ұғым бар (2.1-сурет)
Бұл үш функционалдық әртүрлі элементті қамтитын көз:
1) жарық қабылдайтын, жарықты сындыратын және жарықты реттейтін құрылғылар орналасқан көз алмасы;
2) қорғаныс құралдары, б.а. көздің сыртқы қабықшалары (склера және қасаң қабық), лакримальды аппарат, қабақтар, кірпіктер, қастар; 3) үш жұп көз бұлшықеттерімен (сыртқы және ішкі тік, жоғарғы және төменгі тік ішек, жоғарғы және төменгі қиғаш) бейнеленген қозғалыс аппараты, олар III (көз қозғалғыш нерв), IV (трохлеарлы нерв) және VI (абдусенс нерві) арқылы нервтендіріледі. ) жұп бас сүйек нервтері.
Құрылымдық және функционалдық сипаттамалары
Рецепторлық (перифериялық) бөлім Көру анализаторы (фоторецепторлар) таяқша және конус нейросенсорлық жасушаларға бөлінеді, олардың сыртқы сегменттері сәйкесінше таяқша тәрізді («таяқшалар») және конус тәрізді («конустар»). Адамда 6-7 млн конус және 110-125 млн таяқша болады.
Көру нервінің көз торынан шығатын жерінде фоторецепторлар болмайды және соқыр нүкте деп аталады. Шұңқыр аймағындағы соқыр нүктенің бүйірінде ең жақсы көру аймағы - негізінен конустардан тұратын макула макула орналасқан. Көз торының шетіне қарай конустардың саны азайып, таяқшалардың саны артады, ал тордың шеткі бөлігінде тек таяқшалар болады.
Конустар мен таяқшалардың қызметтеріндегі айырмашылықтар қос көру құбылысының негізінде жатыр. Таяқшалар – аз жарық жағдайында жарық сәулелерін қабылдайтын рецепторлар, яғни. түссіз немесе ахроматикалық көру. Конустар, керісінше, жарқын жарық жағдайында жұмыс істейді және жарықтың спектрлік қасиеттеріне (түсті немесе хроматикалық көрініс) әртүрлі сезімталдықпен сипатталады. Фоторецепторлардың сезімталдығы өте жоғары, бұл рецепторлардың құрылымдық ерекшеліктеріне және жарық тітіркендіргішінің энергиясын қабылдау негізінде жатқан физика-химиялық процестерге байланысты. Фоторецепторлар оларға 1-2 квант жарық әсер еткенде қозылады деп есептеледі.
Таяқшалар мен конустар екі сегменттен тұрады - сыртқы және ішкі, олар бір-бірімен тар кірпікше арқылы байланысқан. Таяқшалар мен конустар торлы қабықта радиалды бағытталған, ал жарық сезгіш белоктардың молекулалары сыртқы сегменттерде олардың жарық сезгіш топтарының шамамен 90%-ы дискілер жазықтығында жататындай етіп орналасады. сыртқы сегменттер. Егер сәуленің бағыты шыбықтың немесе конустың ұзын осімен сәйкес келсе және олардың сыртқы сегменттерінің дискілеріне перпендикуляр бағытталса, жарық ең үлкен қызықты әсерге ие болады.
Көз торындағы фотохимиялық процестер.Тор қабықтың рецепторлық жасушаларында жарыққа сезімтал пигменттер (күрделі ақуызды заттар) – хромопротеиндер болады, олар жарықта түсі өзгереді. Сыртқы сегменттердің қабықшасындағы таяқшаларда родопсин, конустарда йодопсин және басқа пигменттер болады.
Родопсин мен йодопсин торлы қабықтан (А 1 альдегид витамині) және гликопротеиннен (опсин) тұрады. Олардың фотохимиялық процестерде ұқсастықтары болса да, жұтылу максимумы спектрдің әртүрлі аймақтарында болуымен ерекшеленеді. Құрамында родопсин бар таяқшалардың 500 нм аймағында максималды сіңіру мүмкіндігі бар. Конустардың ішінде жұтылу спектрлері бойынша максимумдары бойынша ерекшеленетін үш түрі бар: кейбіреулерінде максимум спектрдің көк бөлігінде (430-470 нм), басқаларында жасылда (500-530 нм) және басқалары бар. қызыл (620-760 нм) бөлігінде, бұл көру пигменттерінің үш түрінің болуына байланысты. Қызыл конус пигменті йодопсин деп аталады. Тор қабықты әртүрлі кеңістіктік конфигурацияларда (изомериялық формаларда) табуға болады, бірақ олардың тек біреуі ғана, ретинальды 11-CIS изомері барлық белгілі көрнекі пигменттердің хромофор тобы ретінде әрекет етеді. Денедегі торлы қабықтың көзі - каротиноидтар.
Тор қабықтағы фотохимиялық процестер өте үнемді жүреді. Жарық сәуленің әсерінен таяқшалардағы родопсиннің аз ғана бөлігі ғана ыдырайды (шамамен 0,006%).
Қараңғыда пигменттердің ресинтезі жүреді, ол энергияны сіңірумен бірге жүреді. Йодопсиннің азаюы родопсинге қарағанда 530 есе жылдам. Денедегі А дәруменінің деңгейі төмендесе, родопсиннің ресинтезі процестері әлсірейді, бұл түнгі соқырлық деп аталатын ымырт көруінің нашарлауына әкеледі. Тұрақты және біркелкі жарықтандыру кезінде пигменттердің ыдырау жылдамдығы мен ресинтезі арасында тепе-теңдік орнатылады. Көз торына түсетін жарық мөлшері азайған кезде бұл динамикалық тепе-теңдік бұзылып, пигменттердің жоғары концентрациясына қарай ығысады. Бұл фотохимиялық құбылыс қараңғы бейімделудің негізінде жатыр.
Фотохимиялық процестерде фузцині бар эпителийден құралған тордың пигменттік қабаты ерекше маңызға ие. Бұл пигмент жарықты сіңіреді, шағылысу мен шашырауды болдырмайды, бұл анық көрнекі қабылдауға әкеледі. Пигментті жасушалардың процестері таяқшалар мен конустардың жарыққа сезімтал сегменттерін қоршап, фоторецепторлардың алмасуына және көру пигменттерінің синтезіне қатысады.
Көздің фоторецепторларындағы фотохимиялық процестерге байланысты жарық әсер еткенде рецепторлық потенциал пайда болады, бұл рецепторлық мембрананың гиперполяризациясы болып табылады. Бұл ерекшеленетін қасиетікөру рецепторлары, басқа рецепторлардың активтенуі олардың мембранасының деполяризациясы түрінде көрінеді. Көру рецепторының потенциалының амплитудасы жарық тітіркендіргішінің қарқындылығы артқан сайын артады. Сонымен, толқын ұзындығы 620-760 нм болатын қызыл сәуленің әсерінен рецепторлық потенциал тордың орталық бөлігінің фоторецепторларында, ал көк (430-470 нм) шеткі бөлігінде айқынырақ болады.
Фоторецепторлардың синаптикалық ұштары биполярлы торлы нейрондарға жақындайды. Бұл жағдайда фовеаның фоторецепторлары тек бір биполярға қосылады.
Сымдар бөлімі.Көру анализаторының өткізгіштік бөлімінің бірінші нейроны көз торының биполярлы жасушаларымен бейнеленген (2.2-сурет).

Күріш. 2.2
Әрекет потенциалдары рецепторлық және көлденең NS ұқсас биполярлы жасушаларда пайда болады деп есептеледі. Кейбір биполярларда жарықты қосу және өшіру кезінде баяу, ұзақ мерзімді деполяризация, ал басқаларында жарықты қосу және өшіру кезінде гиперполяризация, ал сөнген кезде деполяризация жүреді.
Биполярлы жасушалардың аксондары өз кезегінде ганглиондық жасушаларға (екінші нейрон) жиналады. Нәтижесінде әрбір ганглиондық жасуша үшін 140-қа жуық таяқша және 6 конус жиналуы мүмкін, ал макулаға жақын болған сайын, фоторецепторлар бір жасушаға жақындайды. Макула аймағында конвергенция дерлік жоқ және конустардың саны биполярлы және ганглиондық жасушалардың санына тең. Бұл тордың орталық бөліктеріндегі жоғары көру өткірлігін түсіндіреді.
Көз торының перифериясы аз жарыққа өте сезімтал. Бұл, шамасы, 600-ге дейін таяқшаның биполярлы жасушалар арқылы бір ганглиондық жасушаға біріктірілуіне байланысты. Нәтижесінде көптеген таяқшалардың сигналдары жинақталады және бұл жасушалардың неғұрлым қарқынды стимуляциясын тудырады.
Ганглиондық жасушаларда, тіпті толық қараңғылықта да, импульстар тізбегі секундына 5 жиілікпен өздігінен пайда болады. Бұл импульс жалғыз оптикалық талшықтарды немесе жалғыз ганглионды жасушаларды микроэлектродтық зерттеу арқылы анықталады және қараңғыда ол «көздің өз жарығы» ретінде қабылданады.
Кейбір ганглиондық жасушаларда фондық разрядтар жарық қосулы кезде (қосу режимінде), басқаларында - жарық өшірілгенде (өшірулі) жиілік артады. Ганглиондық жасушаның реакциясын жарықтың спектрлік құрамымен де анықтауға болады.
Тор қабықта вертикальдан басқа бүйірлік байланыстар да болады. Рецепторлардың бүйірлік өзара әрекеттесуін көлденең жасушалар жүзеге асырады. Биполярлы және ганглионды жасушалар дендриттердің коллатеральдары мен жасушалардың аксондарымен, сондай-ақ амакриндік жасушалардың көмегімен түзілген көптеген бүйірлік байланыстардың арқасында бір-бірімен әрекеттеседі.
Тор қабықтың көлденең жасушалары фоторецепторлар мен биполярлар арасындағы импульстардың берілуін реттеуді, түсті қабылдауды реттеуді және көздің әртүрлі жарық деңгейіне бейімделуін қамтамасыз етеді. Жарықтандырудың бүкіл кезеңінде көлденең жасушалар оң потенциалды тудырады - S-потенциал (ағылшын тілінен баяу) деп аталатын баяу гиперполяризация. Жарық ынталандыруын қабылдау сипаты бойынша көлденең жасушалар екі түрге бөлінеді:
1) L-типі, онда S-потенциал көрінетін жарықтың кез келген толқынының әсерінен пайда болады;
2) Потенциалды ауытқу белгісі толқын ұзындығына байланысты болатын С-типті немесе «түс» түрі. Осылайша, қызыл жарық олардың деполяризациясын тудыруы мүмкін, ал көк жарық гиперполяризацияны тудыруы мүмкін.
Көлденең ұяшық сигналдары электротоникалық түрде беріледі деп есептеледі.
Көлденең, сондай-ақ амакринді жасушалар ингибиторлық нейрондар деп аталады, өйткені олар биполярлы немесе ганглионды жасушалар арасында бүйірлік тежелуді қамтамасыз етеді.
Бір ганглиондық жасушаға өз сигналдарын жіберетін фоторецепторлар жинағы оның рецептивті өрісін құрайды. Макуланың жанында бұл өрістердің диаметрі 7-200 нм, ал периферияда - 400-700 нм, яғни. Тор қабықтың ортасында рецептивті өрістер кішкентай, ал тордың шеткі бөлігінде диаметрі әлдеқайда үлкен. Тор қабықтың рецептивті өрістері дөңгелек пішінді, концентрлі түрде салынған, олардың әрқайсысында қозу орталығы және сақина түріндегі тежегіш перифериялық аймақ бар. Орталығы қосулы (орталық жарықтандырылғанда қозғалады) және орталықтан тыс (орталық қараңғыланған кезде қозды) қабылдағыш өрістер бар. Тежегіш шекара, қазіргі уақытта болжанғандай, бүйірлік тежелу механизміне сәйкес көлденең торлы жасушалармен қалыптасады, яғни. Қабылдау өрісінің орталығы неғұрлым қоздырылған болса, соғұрлым оның периферияға тежеу әсері күшейеді. Ганглиондық жасушалардың рецептивті өрістерінің (RF) осы түрлерінің арқасында (орталықтары бар және орталықтан тыс) көру аймағындағы ашық және күңгірт нысандар тор қабық деңгейінде анықталады.
Жануарлардың түсті көру қабілеті болса, торлы қабық жасушаларының RP-нің түс-қарсылас ұйымы оқшауланған. Бұл ұйым белгілі бір ганглиондық жасушаның әртүрлі спектрлік сезімталдығы бар конустардан қоздырғыш және ингибиторлық сигналдарды алуынан тұрады. Мысалы, егер «қызыл» конустар берілген ганглиондық жасушаға қоздырғыш әсер етсе, онда «көк» конустар оны тежейді. Конустардың әртүрлі кластарынан қоздырғыш және ингибиторлық кірістердің әртүрлі комбинациялары табылды. Түсі қарама-қарсы ганглиондық жасушалардың айтарлықтай бөлігі конустардың барлық үш түрімен байланысты. РЖ-ның осылай ұйымдастырылуының арқасында жекелеген ганглиондық жасушалар белгілі бір спектрлік құрамды жарықтандыру үшін селективті болады. Сонымен, егер қозу «қызыл» конустардан пайда болса, онда көк және жасыл сезімтал конустардың қозуы бұл жасушалардың тежелуіне әкеледі, ал егер ганглионды жасуша көк-сезімтал конустармен қозса, онда жасыл және қызыл конустармен тежеледі. -сезімталдар және т.б.

Күріш. 2.3
Қабылдау өрісінің орталығы мен перифериясы спектрдің қарама-қарсы ұштарында максималды сезімталдыққа ие. Сонымен, егер қабылдау өрісінің орталығы қызыл жарықтың қосылуына белсенділіктің өзгеруімен жауап берсе, онда периферия көк түстің қосылуына ұқсас реакциямен жауап береді. Бірқатар ретинальды ганглион жасушаларында бағытталған сезімталдық бар. Ол тітіркендіргіш бір бағытта қозғалғанда (оңтайлы) ганглиондық жасуша белсендірілгенімен, тітіркендіргіш басқа бағытта қозғалғанда реакция болмайтындығынан көрінеді. Бұл жасушалардың әртүрлі бағыттағы қозғалысқа реакцияларының селективтілігін ұзартылған процестері бар көлденең жасушалар (теледендриттер) жасайды деп болжанады, олардың көмегімен ганглиондық жасушалар бағытталған түрде тежеледі. Конвергенция және бүйірлік өзара әрекеттесулердің арқасында көрші ганглион жасушаларының рецептивті өрістері қабаттасады. Бұл жарық әсерінің әсерлерін және көз торында өзара тежегіш қатынастардың пайда болуын қорытындылауға мүмкіндік береді.
Тор қабықтағы электрлік құбылыстар.Көру анализаторының рецепторлық бөлімі локализацияланған және өткізгіш бөлімі басталатын көздің торлы қабығында жарықтың әсеріне жауап ретінде күрделі электрохимиялық процестер жүреді, оны жалпы жауап түрінде жазуға болады - электроретинограмма ( ERG) (2.3-сурет).
ERG жарық тітіркендіргішінің түсі, қарқындылығы және әсер ету ұзақтығы сияқты қасиеттерін көрсетеді. ERG бүкіл көзден немесе тікелей тор қабықтан жазылуы мүмкін. Оны алу үшін бір электродты қасаң қабықтың бетіне қояды, ал екіншісін көздің жанындағы бет терісіне немесе құлақшаға жағылады.
Көзді жарықтандыру кезінде жазылған ERG-де бірнеше тән толқындар ерекшеленеді. Бірінші теріс толқын а - бұл фоторецепторлар мен көлденең жасушалардың қозуын көрсететін шағын амплитудалы электрлік тербеліс. Ол тез арада биполярлы және амакринді жасушалардың қозуы нәтижесінде пайда болатын тік өсетін оң b толқынына айналады. b толқынынан кейін баяу электропозитивті с толқыны байқалады - пигменттік эпителий жасушаларының қозуының нәтижесі. Жарықтың тітіркенуінің тоқтау сәті электропозитивті толқынның пайда болуымен байланысты d.
ERG индикаторлары көз аурулары клиникасында көз торының зақымдалуымен байланысты әртүрлі көз ауруларын емдеуді диагностикалау және бақылау үшін кеңінен қолданылады.
Тор қабықтан басталатын өткізгіш бөлім (бірінші нейрон – биполярлы, екінші нейрон – ганглиондық жасушалар) анатомиялық түрде көру нервтерімен және олардың талшықтары ішінара қиылысып өткеннен кейін көру жолдарымен бейнеленген. Әрбір көру жолында сол жақтың тор қабығының ішкі (мұрын) бетінен және екінші көздің тор қабығының сыртқы жартысынан шығатын жүйке талшықтары бар. Оптикалық жолдың талшықтары көрнекі таламусқа (таламустың өзі), метаталамусқа (сыртқы геникулярлық дене) және жастықшаның ядроларына бағытталған. Мұнда көру анализаторының үшінші нейроны орналасқан. Олардан көру жүйке талшықтары ми қыртысына жіберіледі.
Сыртқы (немесе бүйірлік) геникулярлы денелер, торлы қабықтан талшықтар келетін жерде рецептивті өрістер болады, олар да дөңгелек пішінді, бірақ көлемі жағынан торлы қабықтан кішірек. Мұндағы нейрондардың жауаптары фазалық сипатта, бірақ көз торына қарағанда айқынырақ.
Сыртқы геникулярлық денелер деңгейінде көздің тор қабығынан келетін афферентті сигналдардың көру анализаторының кортикальды бөлігінің аймағынан келетін эфферентті сигналдармен әрекеттесу процесі жүреді. Бұл жерде ретикулярлық формацияның қатысуымен есту және басқа да сенсорлық жүйелермен өзара әрекеттесу жүреді, бұл сенсорлық сигналдың ең маңызды компоненттерін бөліп көрсету арқылы таңдамалы көрнекі зейін процестерін қамтамасыз етеді.
Орталық,немесе қыртыстық, бөлімвизуалды анализатор желке бөлігінде (Бродман бойынша 17, 18, 19 өрістер) немесе VI, V2, V3 (қабылданған номенклатура бойынша) орналасқан. Бастапқы проекциялық аймақ (17 өріс) мамандандырылған, бірақ сетчаткаға және сыртқы геникулярлық органдарға қарағанда ақпаратты өңдеуді жүзеге асырады деп саналады. Көру қыртысындағы шағын өлшемді нейрондардың рецептивті өрістері дөңгелек емес, ұзартылған, дерлік төртбұрышты пішіндерге ие. Осымен қатар детекторлық типтегі күрделі және аса күрделі қабылдағыш өрістері бар. Бұл мүмкіндік тұтас кескіннен әр түрлі орналасуы мен бағдары бар сызықтардың жекелеген бөліктерін ғана оқшаулауға мүмкіндік береді және осы фрагменттерге таңдамалы түрде жауап беру мүмкіндігі көрінеді.
Кортекстің әрбір аймағында нейрондар шоғырланған, олар тереңдіктегі барлық қабаттар арқылы тігінен өтетін бағанды құрайды және ұқсас функцияны орындайтын нейрондардың функционалды бірігуі орын алады. Көру объектілерінің әртүрлі қасиеттері (түсі, пішіні, қозғалысы) көру қыртысының әртүрлі бөліктерінде параллельді түрде өңделеді.
Көру қыртысында функционалдық жағынан әртүрлі жасушалар топтары бар - қарапайым және күрделі.
Қарапайым жасушалар қозу және тежеу аймақтарынан тұратын рецептивті өрісті жасайды. Мұны жасушаның кішкене жарық нүктесіне реакциясын зерттеу арқылы анықтауға болады. Күрделі жасушаның рецептивтік өрісінің құрылымын бұлай орнату мүмкін емес. Бұл ұяшықтар көру аймағындағы сызықтардың бұрышын, көлбеуін және қозғалысын детекторлар болып табылады.
Бір бағанда қарапайым және күрделі ұяшықтар болуы мүмкін. Көру қыртысының III және IV қабаттарында таламус талшықтары аяқталатын жерде қарапайым жасушалар табылды. Күрделі жасушалар 17-ші өрістің беткейлік қабаттарында, 18 және 19-ші өрістерде орналасқан, қарапайым жасушалар күрделі және суперкомплексті жасушаларда орналасқан;
Көру қыртысында кейбір нейрондар «қарапайым» немесе концентрлі түсті қарсылас рецептивті өрістерді (IV қабат) құрайды. РЖ-ның түстің оппозициялық болуы орталықта орналасқан нейронның бір түске қозумен әрекеттесуі және басқа түспен қоздырғанда тежелуінен көрінеді. Кейбір нейрондар қызыл жарыққа ашық реакциямен және жасыл жарыққа ofT-жауаппен әрекет етеді, ал басқалары керісінше әрекет етеді.
Концентрлік RP бар нейрондарда түсті рецепторлар (конустар) арасындағы қарсыластық қатынастардан басқа, орталық пен периферия арасында антагонистік қатынастар бар, т.б. Қос түсті қарама-қайшылықпен RP пайда болады. Мысалы, егер нейронда RP центріне әсер еткенде қызыл түске қосулы және жасыл түске қарсы реакция пайда болса, онда оның түс таңдау қабілеті сәйкес түстің жарықтығына селективтілікпен біріктіріледі және ол кез келген ұзындықтағы жарық толқындарымен диффузды ынталандыруға жауап бермейді (from - үшін Польша Республикасының орталығы мен перифериясы арасындағы қарсыластық қатынастар үшін).
Қарапайым RP-де екі немесе үш параллель аймақ бөлінеді, олардың арасында қос қарама-қайшылық бар: егер орталық аймақта қызыл жарыққа қосулы және жасылға өшірулі жауап болса, онда шеткі аймақтарға қарсы жауап береді. қызыл және жасылға жауап.
VI өрістен ми қыртысының медиальды уақытша (ортаңғы уақыттық - МТ) аймағы арқылы басқа (дорсальды) арна өтеді. Бұл аймақтағы нейрондардың жауаптарын тіркеу олардың диспропорцияға (бірдей емес), визуалды әлемдегі объектілердің қозғалысының жылдамдығы мен бағытына жоғары селективті екенін және текстуралы фондағы объектілердің қозғалысына жақсы жауап беретінін көрсетті. Жергілікті жою қозғалатын объектілерге жауап беру қабілетін күрт нашарлатады, бірақ біраз уақыттан кейін бұл қабілет қалпына келеді, бұл бұл аймақ көру өрісіндегі қозғалатын объектілерді талдайтын жалғыз аймақ емес екенін көрсетеді. Бірақ мұнымен қатар, 17 (V1) бастапқы көру өрісінің нейрондары бөлген ақпарат одан әрі көру қыртысының қайталама (V2 өріс) және үшінші (V3 өріс) аймақтарына өңдеу үшін беріледі деп болжанады.
Алайда визуалды ақпаратты талдау жолақты (визуалды) кортекс өрістерінде аяқталмайды (V1, V2, V3). V1 өріс жолдарынан (арналардан) көрнекі сигналдарды одан әрі өңдеу жүзеге асырылатын басқа аймақтарға басталатыны анықталды.
Сонымен, егер сіз уақытша және париетальды аймақтардың түйіскен жерінде орналасқан маймылдағы V4 өрісін жойсаңыз, онда түс пен пішінді қабылдау бұзылады. Пішін туралы көрнекі ақпаратты өңдеу де ең алдымен инферотампоральды аймақта болады деп саналады. Бұл аймақ бұзылған кезде қабылдаудың негізгі қасиеттері (көру өткірлігі және жарықты қабылдау) зардап шекпейді, бірақ жоғары деңгейдегі талдау механизмдері сәтсіздікке ұшырайды.
Сонымен, көрнекі сезім жүйесінде нейрондардың рецептивті өрістері деңгейден деңгейге қарай күрделене түседі, ал синаптикалық деңгей неғұрлым жоғары болса, жеке нейрондардың функциялары соғұрлым қатаң шектеледі.
Қазіргі уақытта көру жүйесі ганглиондық жасушалардан бастап, функционалдық жағынан әртүрлі екі бөлікке (магна- және парвоклеткалы) бөлінеді. Бұл бөліну сүтқоректілердің тор қабығында әртүрлі типтегі ганглиондық жасушалардың болуымен байланысты - X, Y, W. Бұл жасушаларда концентрлі рецептивті өрістер бар, ал олардың аксондары көру жүйкелерін құрайды.
Х-жасушаларында RP шағын, олардың аксондары бойынша қозу жылдамдығы 15-25 м/с анық анықталған; Y жасушаларында әлдеқайда үлкен RP орталығы бар және диффузиялық жарық тітіркендіргіштеріне жақсы жауап береді. Өткізгіштік жылдамдығы 35-50 м/с. Тор қабықта Х жасушалары орталық бөлігін алады, ал шетке қарай олардың тығыздығы төмендейді. Y жасушалары тордың барлық бөлігінде біркелкі таралады, сондықтан тордың шеткі бөлігінде Y жасушаларының тығыздығы X жасушаларынан жоғары. Х-жасушалардың RP-нің құрылымдық ерекшеліктері оларды анықтайды жақсырақ реакциявизуалды ынталандырудың қозғалысын баяулатады, ал Y жасушалары жылдам қозғалатын ынталандыруға жақсырақ жауап береді.
Тор қабықта W жасушаларының үлкен тобы да сипатталған. Бұл ең кішкентай ганглиондық жасушалар, олардың аксондары бойынша өткізгіштік жылдамдығы 5-9 м/с; Бұл топтың жасушалары біртекті емес. Олардың ішінде концентрлі және біртекті РТ бар жасушалар және рецептивті өріс арқылы тітіркендіргіштің қозғалысына сезімтал жасушалар бар. Бұл жағдайда жасуша реакциясы қозғалыс бағытына байланысты емес.
X, Y және W жүйелеріне бөліну геникулярлық дене және көру қыртысы деңгейінде жалғасады. Х нейрондары реакцияның фазалық типіне ие (импульстердің қысқа серпіліс түріндегі активтенуі), олардың рецептивті өрістері перифериялық көру өрістерінде көбірек көрінеді, ал олардың реакциясының жасырын кезеңі қысқа. Бұл қасиеттер жиынтығы олардың жылдам өткізгіш афференттермен қозатынын көрсетеді.
X нейрондарында жауаптың өзекті түрі бар (нейрон бірнеше секунд ішінде белсендіріледі), олардың РҚ көру өрісінің орталығында көбірек бейнеленген, ал жасырын кезең ұзағырақ.
Көру қыртысының бастапқы және қайталама аймақтары (Y1 және Y2 өрістері) X- және Y-нейрондарының мазмұны бойынша ерекшеленеді. Мысалы, Y1 өрісінде X және Y типтерінің екеуінен де афферентация бүйірлік геникулярлық денеден келеді, ал Y2 өрісі афферентті тек Y типті жасушалардан алады.
Көру сенсорлық жүйесінің әртүрлі деңгейлерінде сигналдың берілуін зерттеу көру қабығында (желке аймағында) бас терісінің бетінен электродтарды қолданатын адамды алып тастау арқылы жалпы шақырылған потенциалдарды (ЭП) тіркеу арқылы жүзеге асырылады. Жануарларда көру сенсорлық жүйесінің барлық бөліктеріндегі шақырылған белсенділікті бір уақытта зерттеуге болады.
Әртүрлі жағдайларда анық көруді қамтамасыз ететін механизмдер
Бақылаушыдан әртүрлі қашықтықта орналасқан объектілерді қарастырған кезде, Келесі процестер анық көруді қамтамасыз етеді.
1. Конвергенция және дивергенциялық көз қозғалысы,соның арқасында көрнекі осьтер біріктіріледі немесе бөлінеді. Екі көз бір бағытта қозғалса, мұндай қозғалыстар достық деп аталады.
2. Оқушы реакциясыкөз қозғалысымен синхронды түрде пайда болады. Осылайша, көрнекі осьтердің жақындауымен, жақын орналасқан объектілерді қараған кезде, қарашық тарылады, яғни оқушылардың конвергентті реакциясы. Бұл жауап сфералық аберрациядан туындаған кескіннің бұрмалануын азайтуға көмектеседі. Сфералық аберрация көздің сыну ортасының әртүрлі аймақтарда бірдей емес фокустық қашықтыққа ие болуынан туындайды. Оптикалық ось өтетін орталық бөлік шеткі бөлікке қарағанда үлкен фокустық қашықтыққа ие. Сондықтан торлы қабықтағы сурет бұлыңғыр болады. Көз қарашығының диаметрі неғұрлым аз болса, сфералық аберрациядан болатын бұрмалану соғұрлым аз болады. Көз қарашығының конвергентті тарылуы аккомодациялық аппаратты белсендіріп, линзаның сыну күшін арттырады.

Күріш. 2.4 Көздің аккомодация механизмі: а - тыныштық, б - кернеу

Күріш. 2.5
Қарашық сонымен қатар хроматикалық аберрацияны жоюға арналған құрылғы болып табылады, бұл көздің оптикалық аппараты қарапайым линзалар сияқты қысқа толқынды жарықты ұзын толқынды жарыққа қарағанда күштірек сындырады. Осыған сүйене отырып, қызыл нысанды дәлірек фокустау үшін көкке қарағанда көбірек орналастыру дәрежесі қажет. Сондықтан көк түсті заттар қызылға қарағанда алысырақ көрінеді, олар бірдей қашықтықта орналасқан.
3. Аккомодация – әртүрлі қашықтықтағы объектілердің анық көрінуін қамтамасыз ететін негізгі механизм және алыс немесе жақын объектілерден бейнені көздің торлы қабығына фокустау үшін келеді. Аккомодацияның негізгі механизмі – көз линзасының қисаюының еріксіз өзгеруі (2.4-сурет).
Линзаның, әсіресе алдыңғы бетінің қисаюының өзгеруіне байланысты оның сыну күші 10-14 диоптр шегінде өзгеруі мүмкін. Линза капсуламен қоршалған, ол шеттерінде (линзаның экваторы бойымен) линзаны бекітетін байламға (Зин байламы) өтеді, өз кезегінде кірпікшелі (цилиарлы) бұлшықет талшықтарымен байланысады. Кірпікшелі бұлшықет жиырылғанда зин зонулаларының кернеуі төмендейді, ал линза өзінің серпімділігіне байланысты дөңес болады. Көздің сыну күші артып, көз жақын маңдағы заттарды көруге бейімделеді. Адам алысқа қараған кезде, Зинн байламы шиеленіскен күйде болады, бұл линза қапшығының созылуына және оның қалыңдауына әкеледі. Кірпікшелі бұлшықет симпатикалық және парасимпатикалық нервтермен нервтенеді. Окуломоторлы нервтің парасимпатикалық талшықтары арқылы келетін импульс бұлшықеттің жиырылуын тудырады. Жоғарғы мойын ганглионынан таралатын симпатикалық талшықтар оның босаңсуын тудырады. Кірпікшелі бұлшықеттің жиырылу және босаңсу дәрежесінің өзгеруі көз торының қозуымен байланысты және оған бас миының қыртысы әсер етеді. Көздің сыну күші диоптриямен (D) өрнектеледі. Ауадағы негізгі фокустық арақашықтығы 1 м болатын линзаның сыну қабілетіне бір диоптр сәйкес келеді. Аккомодация құбылысынсыз көздің сыну күші 58-60 D және оны көздің сынуы деп атайды.
Көздің қалыпты сынуы кезінде алыстағы заттардан түсетін сәулелер көздің жарық сыну жүйесінен өткеннен кейін орталық шұңқырдағы торлы қабықтың фокусында шоғырланады. Көздің қалыпты сынуы эмметропия деп аталады, ал мұндай көз эмметропиялық деп аталады. Қалыпты сынумен қатар оның ауытқулары байқалады.
Миопия (миопия) – заттың сәулелері жарықты сындыратын аппарат арқылы өткеннен кейін көз торына емес, оның алдына шоғырланатын сыну қателігінің бір түрі. Бұл көздің үлкен сыну қабілетіне немесе көз алмасының үлкен ұзындығына байланысты болуы мүмкін. Миопиялық адам аккомодациясыз жақын объектілерді көреді, ал алыстағы заттарды анық емес және бұлыңғыр көреді. Түзету үшін екі жақты линзалары бар көзілдірік қолданылады.
Гиперметропия (алыстан көру) – көздің әлсіз сыну қабілетіне немесе көз алмасының қысқа ұзындығына байланысты алыстағы заттардан түсетін сәулелер тордың артына шоғырланатын сыну ақауының бір түрі. Алыстан көретін көз аккомодация кернеуі бар алыстағы объектілерді де көреді, нәтижесінде аккомодативті бұлшықеттердің гипертрофиясы дамиды. Түзету үшін биконвекс линзалар қолданылады.
Астигматизм – әртүрлі меридиандардағы (жазықтықтардағы) көздің қабығы мен линзасының әртүрлі қисаюына байланысты сәулелер бір нүктеге, фокусқа (грек тілінен аударғанда stigme – нүкте) жинала алмайтын сыну ақауының түрі. Астигматизммен объектілер тегістелген немесе ұзартылған болып көрінеді, оны түзету сфероцилиндрлік линзалар арқылы жүзеге асырылады.
Айта кету керек, көздің жарық сыну жүйесіне сонымен қатар: қасаң қабық, көздің алдыңғы камерасының юморы, линза және шыны тәрізді дене кіреді. Бірақ олардың сыну күші линзадан айырмашылығы реттелмейді және аккомодацияға қатыспайды. Сәулелер көздің сыну жүйесінен өткеннен кейін көздің торлы қабығында нақты, қысқартылған және төңкерілген кескін алынады. Бірақ индивидуалды даму процесінде көру анализаторының түйсіктерін моторлы, тері, вестибулярлық және басқа анализаторлардың түйсіктерімен салыстыру, жоғарыда атап өткендей, адамның сыртқы дүниені шын мәнінде сол күйінде қабылдауына әкеледі. .
Бинокулярлық көру (екі көзбен көру) әртүрлі қашықтықтағы объектілерді қабылдауда және оларға дейінгі қашықтықты анықтауда маңызды рөл атқарады, монокулярлық көрумен салыстырғанда кеңістіктің тереңдігінің айқын сезімін береді, яғни. бір көзбен көру. Объектіні екі көзбен қараған кезде оның бейнесі екі көздің тор қабығындағы симметриялы (бірдей) нүктелерге түсуі мүмкін, олардан қозулар анализатордың қыртыстық ұшында біріктіріліп, бір бейнені береді. Егер объектінің кескіні көз торының бірдей емес (алшақ) аймақтарына түссе, онда бөлінген кескін пайда болады. Кеңістікті визуалды талдау процесі бинокулярлық көрудің болуына ғана байланысты емес, бұл жерде көрнекі және моторлы анализаторлар арасында дамитын шартты рефлекторлық әрекеттесулер маңызды рөл атқарады. Кері байланыс принципі бойынша басқарылатын конвергентті көз қозғалысы мен аккомодация процесі белгілі бір мәнге ие. Кеңістікті тұтастай қабылдау анықтамамен байланысты кеңістіктік қатынастаркөрінетін объектілер – олардың көлемі, пішіні, анализатордың әртүрлі бөлімдерінің өзара әрекеттесуімен қамтамасыз етілетін бір-бірімен байланысы; Бұл ретте жинақталған тәжірибе маңызды рөл атқарады.
Объектілер қозғалғандаКелесі факторлар анық көруді қамтамасыз етеді:
1) көз-қозғалыс бұлшықеттерінің конъюгиялық белсенділігі есебінен жүзеге асырылатын объектінің қозғалыс жылдамдығымен көздің жоғары, төмен, солға немесе оңға ерікті қозғалыстары;
2) көру аймағының жаңа бөлігінде зат пайда болған кезде бекіту рефлексі іске қосылады - көздің жылдам еріксіз қозғалысы, көздің торлы қабығындағы заттың кескінінің орталық шұңқырмен сәйкес келуін қамтамасыз етеді. Қозғалатын нысанды қадағалау кезінде көздің баяу қозғалысы пайда болады - қадағалау қозғалысы.
Қозғалмайтын затқа қараған кездеКөздің анық көруін қамтамасыз ету үшін көздер шағын еріксіз қозғалыстардың үш түрін жасайды: тремор - көздің аз амплитудасы мен жиілігімен дірілдеуі, дрейф - көздің айтарлықтай маңызды қашықтыққа баяу ығысуы және секіру (сырттау) - көздің жылдам қозғалысы . Сондай-ақ саккадтық қозғалыстар (саккадалар) бар - екі көздің жоғары жылдамдықпен орындалатын достық қимылдары. Суреттерді оқу және қарау кезінде, визуалды кеңістіктің зерттелетін нүктелері бақылаушыдан және басқа объектілерден бірдей қашықтықта болған кезде сакадтар байқалады. Егер бұл көз қимылдары бітеліп қалса, онда торлы қабық рецепторларының бейімделуіне байланысты бізді қоршаған дүние бақадағыдай айыру қиынға соғады. Бақаның көздері қозғалмайды, сондықтан ол тек қозғалатын заттарды, мысалы, көбелектерді ажырата алады. Сондықтан бақа үнемі тілін шығарып салатын жыланға жақындайды. Қозғалмай қалған бақа жыланды ажыратпай, қозғалатын тілін ұшатын көбелек деп қателеседі.
Өзгеретін жарық жағдайындаайқын көру қарашық рефлексімен, қараңғылық пен жарықтық бейімделумен қамтамасыз етіледі.
Оқушыкөз торына әсер ететін жарық ағынының қарқындылығын оның диаметрін өзгерту арқылы реттейді. Қарашық ені 1,5-тен 8,0 мм-ге дейін өзгеруі мүмкін. Көз қарашығының тарылуы (миоз) жарықтандырудың жоғарылауымен, сондай-ақ жақын маңдағы затты тексергенде және ұйқы кезінде пайда болады. Көз қарашығының кеңеюі (мидриаз) жарықтандырудың төмендеуімен, сондай-ақ рецепторлардың, кез келген афферентті нервтердің қозуымен, симпатикалық жүйке жүйесінің тонусының жоғарылауымен байланысты кернеудің эмоционалдық реакцияларымен (ауырсыну, ашулану, қорқыныш, қуаныш және т.б.), психикалық қозумен (психоз, истерия және т.б.), тұншығумен, наркозбен. Қарашық рефлексі жарықтандыру өзгерген кезде көрнекі қабылдауды жақсартса да (ол қараңғыда кеңейеді, бұл торға түсетін жарық ағынын арттырады, жарықта тарылады), негізгі механизм әлі де қараңғы және жарыққа бейімделу болып табылады.
Темпаға бейімделукөру анализаторының сезімталдығының жоғарылауында көрінеді (сенсибилизация), жарыққа бейімделу- көздің жарыққа сезімталдығын төмендету. Жарық және қараңғылық бейімделу механизмдерінің негізіне фотосезімтал пигменттердің бөлінуін (жарықта) және ресинтезін (қараңғыда) қамтамасыз ететін конустар мен таяқшаларда болатын фотохимиялық процестер, сондай-ақ функционалдық қозғалғыштық процестері: қосу болып табылады. және көз торының рецепторлық элементтерінің белсенділігінен. Сонымен қатар, бейімделу белгілі бір нейрондық механизмдермен және, ең алдымен, тордың нейрондық элементтерінде болатын процестермен, атап айтқанда, көлденең және биполярлы жасушалардың қатысуымен фоторецепторларды ганглиондық жасушаларға қосу әдістерімен анықталады. Осылайша, қараңғыда бір биполярлы жасушамен байланысқан рецепторлардың саны көбейеді және олардың көпшілігі ганглиондық жасушаға жиналады. Сонымен бірге әрбір биполярлы және табиғи түрде ганглиондық жасушаның рецептивті өрісі кеңейеді, бұл көру қабылдауды жақсартады. Көлденең жасушалардың қосылуы орталық жүйке жүйесімен реттеледі.
Симпатикалық жүйке жүйесінің тонусының төмендеуі (көздің десимпатизациясы) қараңғы бейімделу жылдамдығын төмендетеді, ал адреналинді енгізу керісінше әсер етеді. Ми діңінің ретикулярлық формациясының тітіркенуі көру нервтерінің талшықтарындағы импульстардың жиілігін арттырады. Орталық жүйке жүйесінің тор қабықтағы бейімделу процестеріне әсері басқа көзге жарық түскенде және дыбыс, иіс сезу немесе дәм сезу тітіркендіргіштерінің әсерінен жарықсыз көздің жарыққа сезімталдығының өзгеруімен де дәлелденеді.
Түс бейімдеу.Ең жылдам және ең драмалық бейімделу (сезімталдықтың төмендеуі) көк-күлгін тітіркендіргіштің әсерінен болады. Қызыл ынталандыру ортаңғы орынды алады.
Үлкен объектілерді және олардың бөлшектерін визуалды қабылдауорталық және перифериялық көру есебінен қамтамасыз етіледі – көру бұрышының өзгеруі. Нысанның ұсақ бөлшектерін ең дәл бағалау, егер кескін тордың орталық шұңқырында локализацияланған макулаға түссе, қамтамасыз етіледі, өйткені бұл жағдайда ең үлкен көру өткірлігі пайда болады. Бұл макула аймағында тек конустардың болуымен түсіндіріледі, олардың өлшемдері ең кішкентай және әрбір конус аздаған нейрондармен байланыста болады, бұл көру өткірлігін арттырады. Көру өткірлігі көз әлі де екі нүктені бөлек көре алатын ең кішкентай көру бұрышымен анықталады. Қалыпты көз 1" көру бұрышында екі жарық нүктені ажырата алады. Мұндай көздің көру өткірлігі бір деп алынады. Көру өткірлігі көздің оптикалық қасиеттеріне, тор қабықтың құрылымдық ерекшеліктеріне және жұмысына байланысты. Көру анализаторының өткізгіш және орталық бөлімдерінің нейрондық механизмдері алфавиттік немесе әртүрлі типтегі фигуралық стандартты кестелердің көмегімен анықталады. көзқарас.
Көру өрісі - тұрақты көзбен көруге болатын кеңістік. Сол және оң көздер үшін бөлек көру өрістері, сондай-ақ екі көз үшін ортақ көру өрісі бар. Адамның көру аймағының мөлшері көз алмасының тереңдігіне және қас жоталары мен мұрынның пішініне байланысты. Көру өрісінің шекаралары көздің көру осінен құралған бұрышпен және көздің түйіндік нүктесі арқылы көздің тор қабығына қарай шеткі көрінетін нүктеге түсірілген сәулемен белгіленеді. Көру өрісі әртүрлі меридиандарда (бағыттарда) бірдей емес. Төмен – 70°, жоғары – 60°, сыртқа – 90°, ішке қарай – 55°. Көз торының шеткі жағында түсті (конусты) қабылдайтын рецепторлардың болмауына байланысты ахроматикалық көру өрісі хроматикалықға қарағанда үлкенірек. Өз кезегінде, түс көру өрісі әртүрлі түстер үшін бірдей емес. Жасыл үшін ең тар көру өрісі, сары үшін, қызыл үшін кеңірек, үшін одан да кеңірек көк түстер. Көру өрісінің өлшемі жарықтандыруға байланысты өзгереді. Ахроматикалық көру өрісі ымыртта ұлғаяды, ал жарықта азаяды. Хроматикалық көру өрісі, керісінше, жарықта ұлғаяды, ымыртта азаяды. Бұл фоторецепторларды мобилизациялау және демобилизациялау процестеріне байланысты (функционалдық мобильділік). Ымырттың көрінісімен жұмыс істейтін таяқшалар санының артуы, яғни. олардың мобилизациясы көрудің ахроматикалық өрісінің ұлғаюына әкеледі, сонымен бірге жұмыс істейтін конустардың санының азаюы (олардың демобилизациясы) көрудің хроматикалық өрісінің төмендеуіне әкеледі (П.Г. Снякин).
Көрнекі анализатордың тетігі де бар жарық толқындарының ұзындығын ажырату -түсті көру.
Түс көру, визуалды контраст және дәйекті кескіндер
Түс көру - визуалды анализатордың түс сезімін қалыптастыру арқылы жарық толқын ұзындығының өзгеруіне жауап беру қабілеті. Электромагниттік сәулеленудің белгілі бір толқын ұзындығы белгілі бір түстің сезіміне сәйкес келеді. Осылайша, қызыл түстің сезімі толқын ұзындығы 620-760 нм, ал күлгін - 390-450 нм болатын жарықтың әрекетіне сәйкес келеді, спектрдің қалған түстері аралық параметрлерге ие. Барлық түстерді араластыру сезім береді ақ. Спектрдің үш негізгі түсін - қызыл, жасыл, көк-күлгін - әртүрлі пропорцияда араластыру нәтижесінде кез келген басқа түстерді қабылдауға болады. Түстерді сезіну жарықтандырумен байланысты. Ол азайған сайын алдымен қызыл түстер, ал көк түстер кейінірек ажыратылуын тоқтатады. Түсті қабылдау негізінен фоторецепторларда болатын процестермен анықталады. Ең үлкен танутүсті қабылдаудың үш компонентті Ломоносов-Юнг-Гельмгольц-Лазарев теориясын қолданады, соған сәйкес көздің тор қабығында қызыл, жасыл және көк-күлгін түстерді бөлек қабылдайтын фоторецепторлардың үш түрі – конустар болады. Әртүрлі конустарды ынталандыру комбинациясы әртүрлі түстер мен реңктердің сезіміне әкеледі. Конустардың үш түрін біркелкі ынталандыру ақ түс сезімін береді. Түс көрудің үш компонентті теориясы Р.Граниттің (1947) электрофизиологиялық зерттеулерінде расталды. Түске сезімтал конустардың үш түрін модуляторлар деп атады, олар жарық жарықтығының өзгеруімен қоздырылды (төртінші түрі) доминанттар деп аталды. Кейіннен микроспектрофотометрияны қолдана отырып, тіпті бір конустың әртүрлі толқын ұзындығының сәулелерін жұта алатынын анықтау мүмкін болды. Бұл әр конуста әртүрлі ұзындықтағы жарық толқындарына сезімтал түрлі пигменттердің болуына байланысты.
Үш компонентті теорияның сенімді дәлелдеріне қарамастан, түстерді көру физиологиясында осы позициялардан түсіндіруге болмайтын фактілер сипатталған. Бұл қарама-қарсы немесе қарама-қарсы түстер теориясын алға тартуға мүмкіндік берді, т.б. Эвальд Херингтің түсті көрудің қарсылас теориясын жасау.
Бұл теорияға сәйкес, көзде және/немесе мида үш қарсылас процесс бар: біреуі қызыл және жасылды сезіну үшін, екіншісі сары және көкті сезіну үшін және үшіншісі алғашқы екі процестен сапалы түрде ерекшеленеді - қара және ақ үшін. Бұл теория визуалды жүйенің келесі бөліктерінде түс туралы ақпараттың берілуін түсіндіру үшін қолданылады: ретинальды ганглион жасушалары, сыртқы геникулярлық денелер, кортикальды көру орталықтары, мұнда түсті қарсылас РП олардың орталығымен және перифериясымен жұмыс істейді.
Осылайша, алынған мәліметтерге сүйене отырып, конустардағы процестер түсті қабылдаудың үш компонентті теориясына көбірек сәйкес келеді деп болжауға болады, ал Герингтің қарама-қарсы түстер теориясы көз торының нейрондық желілері мен оның үстіне жатқан көру орталықтары үшін қолайлы.
Түсті қабылдауда нейрондарда болатын процестер де белгілі бір рөл атқарады. әртүрлі деңгейлервизуалды анализатор (соның ішінде сетчатка), олар түсті қарсылас нейрондар деп аталады. Көзге спектрдің бір бөлігінен сәулелену әсер еткенде, ол қозып, екінші бөлігімен тежеледі. Мұндай нейрондар түс ақпаратын кодтауға қатысады.
Түс көруіндегі ауытқулар байқалады, олар ішінара немесе толық түсті соқырлық ретінде көрінуі мүмкін. Түстерді мүлде ажырата алмайтын адамдарды ахроматтар деп атайды. Жартылай түсті соқырлық ерлердің 8-10% -ында және әйелдердің 0,5% -ында кездеседі. Түс соқырлығы еркектерде жыныстық жұпталмаған Х хромосомасында белгілі бір гендердің болмауымен байланысты деп саналады. Жартылай түс соқырлығының үш түрі бар: протанопия(түс соқырлығы) – ең алдымен қызыл түске соқырлық. Түс соқырлығының бұл түрін алғаш рет 1794 жылы аномалияның осы түрін байқаған физик Дж.Дальтон сипаттаған. Аномалияның бұл түрі бар адамдар «қызыл соқырлар» деп аталады; дейтеранопия- жасыл түсті қабылдаудың төмендеуі. Мұндай адамдарды «жасыл соқырлар» деп атайды; тританопия- сирек кездесетін аномалия. Алайда адамдар көк және күлгін түстерді қабылдамайды, олар «күлгін-соқыр» деп аталады.
Түс көрудің үш компонентті теориясы тұрғысынан аномалияның әрбір түрі үш конус түсті қабылдау субстраттарының біреуінің болмауының нәтижесі болып табылады. Түс көруінің бұзылуын диагностикалау үшін олар Э.Б.Рабкиннің түс кестелерін, сондай-ақ арнайы құрылғыларды пайдаланады. аномалоскоптар.Түрлі түсті көру ауытқуларын анықтау үлкен құндылықадамның әртүрлі жұмыс түрлеріне (жүргізуші, ұшқыш, суретші және т.б.) кәсіби жарамдылығын анықтау кезінде.
Түсті қабылдау қабілетінде көрінетін жарықтың толқын ұзындығын бағалау қабілеті адам өмірінде маңызды рөл атқарады, эмоционалдық салаға және әртүрлі дене жүйелерінің қызметіне әсер етеді. Қызыл түс жылу сезімін тудырады, психикаға ынталандырушы әсер етеді, эмоцияларды күшейтеді, бірақ тез шаршайды, бұлшықет кернеуіне, қан қысымының жоғарылауына және тыныс алудың жоғарылауына әкеледі. Апельсин түсі көңілділік пен әл-ауқат сезімін тудырады және ас қорытуды жақсартады. Сары түс жақсы, көтеріңкі көңіл-күй тудырады, көруді ынталандырады және жүйке жүйесі. Бұл ең «қызықты» түс. Жасыл түс сергітетін және тыныштандыратын әсерге ие, ұйқысыздыққа, шаршауға пайдалы, қан қысымын, дененің жалпы тонусын төмендетеді және адам үшін ең қолайлы. Көк түс салқындық сезімін тудырады және жүйке жүйесіне тыныштандыратын әсер етеді және жасыл түске қарағанда күшті (көк түс әсіресе жүйке қозғыштығы жоғары адамдар үшін қолайлы), жасыл түске қарағанда қан қысымы мен бұлшықет тонусын төмендетеді. Күлгін түс соншалықты тыныштандырмайды, өйткені ол психиканы босаңсытады. Адам психикасы қызылдан күлгінге дейінгі спектрді қадағалай отырып, эмоциялардың бүкіл гаммасын басып өтетін сияқты. Бұл анықтау үшін Люшер сынамасын қолданудың негізі болып табылады эмоционалдық күйдене.
Көрнекі контрасттар және дәйекті кескіндер.Тітіркену тоқтағаннан кейін көрнекі сезімдер жалғасуы мүмкін. Бұл құбылыс тізбектелген бейнелер деп аталады. Көрнекі контрасттар - бұл қоршаған жарық немесе түсті фонға байланысты ынталандыруды қабылдаудың өзгертілген. Жарық және түсті визуалды контраст деген ұғымдар бар. Қарама-қарсылық құбылысы бір мезгілде немесе бірізді екі түйсіктің нақты айырмашылығын әсірелеуде көрінуі мүмкін, сондықтан бір мезгілде және ретті қарама-қайшылықтар арасында айырмашылық жасалады. Ақ фонда сұр жолақ күңгірт фонда орналасқан сол жолаққа қарағанда күңгірт болып көрінеді. Бұл бір мезгілде жарық контрастының мысалы. Сұр түсті қызыл фонда қарасақ, ол жасылдау болып көрінеді, ал көк фонда сұр түске қарасақ, сарғыш болып көрінеді. Бұл бір мезгілде түс контрастының құбылысы. Тұрақты түсті контраст - бұл ақ фонда қараған кезде түс сезімінің өзгеруі. Сонымен, егер сіз қызыл түске боялған бетке ұзақ қарап, содан кейін көзіңізді ақ түске бұрсаңыз, ол жасыл реңкке ие болады. Көрнекі контрасттың себебі тордың фоторецепторында және нейрондық аппаратында болатын процестер болып табылады. Негізі тордың әртүрлі рецептивті өрістеріне жататын жасушалардың өзара тежелуі және олардың анализаторлардың кортикальды бөлігіндегі проекциялары.
- Көру анатомиясы
Көру анатомиясы
Көру құбылысы
Ғалымдар түсіндіргенде көру құбылысы , олар көбінесе көзді камерамен салыстырады. Жарық, құрылғының линзаларындағы сияқты, көзге кішкентай тесік арқылы кіреді - иристің ортасында орналасқан қарашық. Қарашық кеңірек немесе тар болуы мүмкін: осылайша түсетін жарық мөлшері реттеледі. Әрі қарай, жарық көздің артқы қабырғасына - торға бағытталады, нәтижесінде мида белгілі бір сурет (бейне, сурет) пайда болады. Сол сияқты, жарық камераның артына түскенде, сурет пленкаға түсіріледі.
Біздің көзқарасымыз қалай жұмыс істейтінін егжей-тегжейлі қарастырайық.
Біріншіден, көздің көрінетін бөліктері, олар тиесілі, жарық алады. Ирис(«енгізу») және склера(көздің ағы). Көз қарашығынан өткеннен кейін жарық фокустаушы линзаға түседі ( линза) адамның көзі. Жарықтың әсерінен көздің қарашығы адамның күш-жігерінсіз немесе бақылаусыз жиырылады. Бұл ирис бұлшықеттерінің бірі болғандықтан болады сфинктері- жарыққа сезімтал және оған кеңею арқылы әрекет етеді. Қарашықтардың тарылуы миымызды автоматты түрде басқаруға байланысты пайда болады. Заманауи фокустау камералары бірдей нәрсені жасайды: фотоэлектрлік «көз» линзаның артындағы кіретін тесіктің диаметрін реттейді, осылайша түсетін жарық мөлшерін мөлшерлейді.
Енді көз линзасының артындағы кеңістікке жүгінейік, онда линза, шыны тәрізді желатинді зат орналасқан ( шыны тәрізді) және соңында - көз торы, оның құрылымына шынайы таңдануды тудыратын орган. Тор қабық көз түбінің кең бетін жабады. Бұл денедегі басқа құрылымдарға ұқсамайтын күрделі құрылымы бар ерекше орган. Көздің торлы қабығы «таяқшалар» және «конустар» деп аталатын жүздеген миллион жарыққа сезімтал жасушалардан тұрады. фокуссыз жарық. Таяқшаларқараңғыда көруге арналған, ал олар айналысқанда біз көрінбейтінін қабылдай аламыз. Фотопленка бұған қабілетті емес. Жартылай қараңғыда түсіруге арналған пленканы пайдалансаңыз, ол ашық жарықта көрінетін суретті түсіре алмайды. Бірақ адам көзінің бір ғана торлы қабығы бар және ол әртүрлі жағдайларда жұмыс істеуге қабілетті. Мүмкін оны көп функциялы фильм деп атауға болады. Конустар, таяқшалардан айырмашылығы, жарықта жақсы жұмыс істейді. Олар айқын фокус пен айқын көруді қамтамасыз ету үшін жарық қажет. Конустардың ең жоғары концентрациясы сетчатканың макула («дақ») деп аталатын аймағында болады. Бұл нүктенің орталық бөлігінде fovea centralis (көз ұясы немесе шұңқыр): дәл осы аймақ ең өткір көруді мүмкін етеді.
Мүйізді қабық, қарашық, линза, шыны тәрізді дене, сондай-ақ көз алмасының өлшемі - белгілі бір құрылымдардан өткен кезде жарықтың фокусталуы осының барлығына байланысты.Жарық фокусының өзгеру процесі сыну деп аталады. Неғұрлым дәлірек бағытталған жарық фовеаға түседі, ал азырақ фокусталған жарық көз торына таралады.
Біздің көзіміз жарық қарқындылығының он миллионға жуық градациясын және түстердің жеті миллионға жуық реңктерін ажырата алады.
Дегенмен, көру анатомиясы мұнымен шектелмейді. Көру үшін адам бір уақытта көзін де, миын да пайдаланады, бұл үшін камерамен қарапайым ұқсастық жеткіліксіз. Әрбір секунд сайын көз миға миллиардқа жуық ақпарат жібереді (біз қабылдайтын барлық ақпараттың 75 пайызынан астамы). Жарықтың бұл бөліктері сіздің санаңызда сіз танитын таңғажайып күрделі бейнелерге айналады. Жарық, осы танылатын бейнелердің пішінін ала отырып, өткен оқиғалар туралы естеліктеріңіз үшін ынталандырушы ретінде пайда болады. Бұл мағынада көру тек пассивті қабылдау ретінде әрекет етеді.
Көретіндердің барлығы дерлік біз көруді үйренгеніміз. Өйткені, біз көз торына түсетін жарықтан ақпаратты қалай алуға болатынын білмей-ақ өмірге келеміз. Нәрестелік шақта біз көрген нәрселер біз үшін ештеңені білдірмейді немесе дерлік ештеңе білдірмейді. Тор қабықшадан жарықпен ынталандырылған импульстар миға енеді, бірақ нәресте үшін олар тек сезім, мағынасы жоқ. Адам есейіп, үйренген сайын бұл сезімдерді түсіндіре бастайды, оларды түсінуге тырысады, олардың нені білдіретінін түсінуге тырысады.
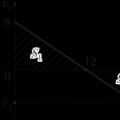 Лездік қозғалыс жылдамдығы
Лездік қозғалыс жылдамдығы Тригонометриялық функциялардың интегралдары
Тригонометриялық функциялардың интегралдары Етістіктің шағы қалай анықталады?
Етістіктің шағы қалай анықталады?