Бір ген бір ферменттің заманауи түсіндірмесі. Бір ген – бір фермент теориясы
4.2.1. «Бір ген, бір фермент» гипотезасы
Алғашқы зерттеу.Гаррод 1902 жылы алкаптонуриядағы генетикалық ақау организмнің гомогентизин қышқылын ыдыра алмауымен байланысты екенін көрсеткеннен кейін, бұл бұзылыстың негізінде жатқан ерекше механизмді түсіндіру маңызды болды. Ол кезде метаболикалық реакциялар ферменттермен катализденетіні белгілі болғандықтан, бұл алкаптонурияға әкелген кейбір ферменттердің бұзылуы деп болжауға болады. Мұндай гипотезаны Дриеш (1896 ж.) талқылады. Оны Халден (1920, қараңыз) және Гаррод (1923) де білдірді. Маңызды кезеңдеріБиохимиялық генетиканың дамуы Кюхн мен Бутенандттың диірмен көбелегіндегі көздің түсін зерттеу жұмыстарынан басталды. Ephestia kuhniellaжәне ұқсас зерттеулер Beadle және Ephrussi бойынша Дрозофила(1936). Бұл жаңашыл жұмыстарда гендік әсер ету механизмдерін түсіндіру үшін бұрын генетикалық әдістермен зерттелген жәндіктердің мутанттары таңдалды. Алайда, бұл әдіс табысқа әкелмеді. Мәселе тым күрделі болып шықты және оны шешу үшін қажет болды:
1) эксперименттік зерттеуге ыңғайлы қарапайым модельді ағзаны таңдау;
2) генетикалық анықталған белгілердің биохимиялық негізін емес, биохимиялық белгілердің генетикалық негізін іздеу. Екі шарт 1941 жылы Бидл мен Татумның жұмысында орындалды (сонымен қатар Beadle, 1945 қараңыз).
Бидл мен Татумның үлгісі. Бұл зерттеушілердің мақаласы былай басталды:
«Физиологиялық генетика тұрғысынан организмнің дамуы мен жұмыс істеуі қандай да бір түрде гендермен басқарылатын күрделі химиялық реакциялар жүйесіне дейін қысқартылуы мүмкін. Бұл гендер ... өздері фермент ретінде әрекет етеді немесе олардың ерекшелігін анықтайды деп болжау әбден қисынды. Генетик-физиологтар әдетте бұрыннан белгілі тұқым қуалайтын белгілердің физиологиялық және биохимиялық негіздерін зерттеуге тырысатыны белгілі. Бұл тәсіл көптеген биоларды құруға мүмкіндік берді химиялық реакцияларарнайы гендермен бақыланады. Мұндай зерттеулер ферменттер мен гендердің бір ретті спецификаға ие екенін көрсетті. Дегенмен, бұл тәсілдің мүмкіндіктері шектеулі. Ең маңызды шектеу - бұл жағдайда зерттеушілер өлімге әкелетін әсері жоқ тұқым қуалайтын белгілерге назар аударады, демек, организмнің өмірі үшін өте маңызды емес реакциялармен байланысты. Екінші қиындық... мәселеге дәстүрлі көзқарас сырттан көрінетін белгілерді қолдануды көздейді. Олардың көпшілігі биохимиялық реакция жүйелеріне негізделген морфологиялық вариациялар болып табылатыны соншалық, оларды талдау өте қиын.
Осыған ұқсас ойлар бізді мынадай қорытындыға әкелді. Даму мен метаболизмді анықтайтын биохимиялық реакцияларды генетикалық бақылаудың жалпы мәселесін зерттеуді қолдану арқылы жүзеге асыру керек. жалпы қабылданғанға қарсы рәсім:Белгілі тұқым қуалайтын белгілердің химиялық негізін анықтауға тырысудың орнына, оны анықтау керек гендер белгілі биохимиялық реакцияларды басқарады ма және қалай.Аскомицеттерге жататын нейроспоралар мұндай тәсілді жүзеге асыруға мүмкіндік беретін қасиеттерге ие және сонымен бірге генетикалық зерттеуге ыңғайлы объект ретінде қызмет етеді. Сондықтан біздің бағдарламамыз осы нақты ағзаны пайдалануға негізделген. Біз рентгендік әсер белгілі бір химиялық реакцияларды басқаратын гендердегі мутацияларды тудырады деп болжадық. Белгілі бір ортада тіршілік ету үшін организм қандай да бір химиялық реакция жүргізуі керек делік, онда мұндай қабілетсіз мутант бұл жағдайда өмір сүру мүмкін емес болады. Дегенмен, оны генетикалық блокталған реакцияның өмірлік өнімі қосылған ортада өсірген жағдайда сақтауға және зерттеуге болады ».
4 Гендердің әрекеті 9
Күріш. 4.1. Нейроспораның биохимиялық мутанттарын анықтау бойынша эксперимент схемасы Толық ортада рентген сәулелері немесе ультракүлгін сәулелер әсерінен туындаған мутациялар саңырауқұлақтың өсуін бұзбайды. Алайда мутант минималды ортада өспейді. Витаминдерді минималды ортаға қосқанда өсу қабілеті қалпына келеді.Аминқышқылдарын қосқанда өсу болмайды.Осы мәліметтерге сүйене отырып, мутация витамин алмасуын бақылайтын генде болған деп болжауға болады. Келесі қадам қалыпты функцияны қалпына келтіре алатын витаминді анықтау болып табылады.Дәрумендердің биосинтезі реакцияларының арасында генетикалық блок кездеседі.Одан әрі Бидл мен Татум тәжірибелік жобаны сипаттайды (4.1-сурет). Толық ортада агар, бейорганикалық тұздар, уыт сығындысы, ашытқы сығындысы және глюкоза бар. Минималды ортада тек агар, тұздар, биотин және көміртек көзі болды. Толық ортада өскен және минималды ортада өспеген мутанттар барынша егжей-тегжейлі зерттелді. Мутанттардың әрқайсысында синтезі бұзылған қосылысты анықтау үшін толық ортаның жеке компоненттері минималды агарға қосылды.
Осылайша кейбір өсу факторларын синтездей алмайтын штаммдар бөлініп алынды: пиридоксин, тиамин және пара-аминобензой қышқылы. Бұл ақаулар белгілі бір локустардағы мутациялардан туындайтыны дәлелденді. Жұмыс нейроспоралар, бактериялар және ашытқылар бойынша көптеген зерттеулердің негізін қалады, оларда метаболизмнің жекелеген кезеңдері мен спецификалық фермент бұзылыстарына жауап беретін «генетикалық блоктардың» сәйкестігі анықталды. Бұл тәсіл тез арада зерттеушілерге метаболизм жолдарын ашуға мүмкіндік беретін құралға айналды.
«Бір ген – бір фермент» гипотезасы күшті эксперименталды растауға ие болды. Кейінгі онжылдықтардағы жұмыс көрсеткендей, бұл таңқаларлықтай жемісті болды. Ақаулы ферменттерді және олардың қалыпты нұсқаларын талдау көп ұзамай ферменттің функциясының өзгеруіне әкелетін генетикалық бұзылулар класын анықтауға мүмкіндік берді, дегенмен белоктың өзі әлі де анықталып, иммунологиялық қасиеттерін сақтаған. Басқа жағдайларда ферменттердің белсенділігі үшін оңтайлы температура өзгерді. Кейбір нұсқаларды жалпы реттеу механизміне әсер ететін мутациямен және соның нәтижесінде ферменттердің бүкіл тобының белсенділігін өзгертумен түсіндіруге болады. Осыған ұқсас зерттеулер бактериялардағы гендердің белсенділігін реттеу концепциясын жасауға әкелді, оған оперон ұғымы кірді.
10 4. Гендердің әрекеті
Адамдардағы ферментативті бұзылулардың алғашқы мысалдары.Ферментативті бұзылыстарды көрсетуге болатын адамның бірінші тұқым қуалайтын ауруы тұқым қуалаудың рецессивті режимі бар метгемоглобинемия болды (Гибсон және Харрисон, 1947; Гибсон, 1948) (25080). Бұл жағдайда зақымдалған фермент NADH - тәуелді метгемоглобин редуктаза болып табылады. Зат алмасу ақауларымен байланысты адам ауруларының тобын жүйелі түрде зерттеуге алғашқы әрекет 1951 жылы жасалды. Корейлер гликогенді сақтау ауруын зерттеуде Гиерке ауруы (23220) диагнозы қойылған патологиялық жағдайдың он жағдайының сегізінде бауырдың гликоген құрылымы қалыпты, ал екі жағдайда айқын ауытқушылық болғанын көрсетті. Сондай-ақ артық жиналатын бауырдағы гликоген тікелей қантқа айналмайтыны анық болды, өйткені пациенттер гипогликемияға бейім болды. Бауырда глюкоза түзу үшін гликогенді ыдырату үшін көптеген ферменттер қажет. Олардың екеуі, амило-1,6-глюкозидаза және глюкоза 6-фосфатаза ферменттік жүйенің мүмкін ақаулы элементтері ретінде зерттеу үшін таңдалды. Глюкоза-6фосфаттан фосфаттың бөлінуі әртүрлі рН мәндерінде бауыр гомогенаттарында өлшенді. Нәтижелер суретте көрсетілген. 4.2. Қалыпты бауырда рН 6-7 кезінде оңтайлы жоғары белсенділік анықталды. Бауыр циррозындағы ауыр дисфункция белсенділіктің аздап төмендеуімен байланысты. Екінші жағынан, өліммен аяқталатын Гиерке ауруы жағдайында ферменттің белсенділігін мүлдем анықтау мүмкін болмады; ұқсас нәтиже екінші ұқсас науқасты тексергенде алынған. Симптомдары онша ауыр емес екі пациентте белсенділіктің айтарлықтай төмендеуі байқалды.
Бұл өлімге әкелетін Гиерке ауруында глюкоза-6-фосфатазаның ақауы бар деген қорытынды жасалды. Алайда, ең жеңіл жағдайларда бұл ферменттің белсенділігі бауыр циррозындағыдан төмен болған жоқ, тек екі науқаста ол сәл төмен болды (4.2-сурет).
Корейлердің пікірінше, бұлшықет тінінде гликогеннің қалыптан тыс жиналуын глюкоза-6-фосфатаза жетіспеушілігімен байланыстыруға болмайды, өйткені бұл фермент бұлшықеттерде жоқ және қалыпты. Олар бұлшықет гликогенозының ықтимал түсіндірмесі ретінде амило-1,6-глюкозидаза белсенділігінің бұзылуын ұсынды. Бұл болжам көп ұзамай расталды: Форбс жүрек және қаңқа бұлшықеттерін қамтитын гликогенді сақтау ауруының клиникалық маңызды жағдайларының бірінде мұндай ақауды тапты. Енді біз
4. Гендердің әрекеті 11
Гликогенді сақтау ауруында көптеген ферментативті ақаулар белгілі.
Бұл аурудың әртүрлі формалары көрініс дәрежесінде біршама ерекшеленсе де, клиникалық тұрғыдан олардың ортақ жақтары көп. Бір қоспағанда, олардың барлығы аутосомды-рецессивті жолмен тұқым қуалайды. Егер ферментативті ақаулар табылмаса, гликогенді сақтау патологиясы ауырлық дәрежесінде, симптоматикалық мәліметтерде және өлім уақытында тән интрафамилиялық корреляциясы бар бір ауру ретінде қарастырылатын еді. Осылайша, біздің алдымызда фенотипті (3.3.5-бөлім) зерттеу негізінде ғана болжауға болатын генетикалық гетерогенділік биохимиялық деңгейде талдау арқылы расталған мысал бар: зерттеу ферментативті белсенділікарнайы гендерді анықтауға мүмкіндік берді.
Кейінгі жылдарда ферментативті ақаулар саласындағы зерттеулердің қарқыны артты және МакКусик өзінің «Адамдағы мендельдік тұқымқуалаушылық» кітабының алтыншы басылымында сипаттайтын 588 анықталған рецессивті аутосомды гендер үшін спецификалық ферментативті ақаулар астам табылды. 170 жағдай. Бұл саладағы жетістіктеріміз молекулалық генетиканың тұжырымдамалары мен әдістерінің дамуына тікелей байланысты.
Адамдардағы ферментативті бұзылыстарды зерттеудің кейбір кезеңдері.Біз осы үздіксіз процестің ең маңызды кезеңдерін ғана ұсынамыз: 1934 жылы Фоллинг фенилкетонурияны ашты.
1941 ж. Бидл мен Татум «бір ген, бір фермент» гипотезасын тұжырымдады 1948 ж. Гибсон адам ауруындағы ферментативті бұзылыстың алғашқы жағдайын сипаттады (рецессивті метгемоглобинемия)
1952 Корейлер Гиерке ауруында глюкоза-6-фосфатаза тапшылығын анықтады.
1953 Джервис фенилкетонурияда фенилаланин гидроксилазасының жоқтығын көрсетті. Бикель фенилаланинді төмен диетаны қолдана отырып, ферментативті бұзылуды жеңілдетудің алғашқы әрекетін хабарлады
1955 Smithies крахмал гель электрофорезі әдісін жасады
1956 Карсон және басқалар индукцияланған гемолитикалық анемия жағдайында глюкоза-6-фосфатдегидрогеназаның (G6PD) ақауын анықтады.
1957 Қалқар және т.б. галактоземиядағы ферменттік тапшылықты сипаттап, адамдар мен бактериялардың ферментативті белсенділігінде бірдей ақауларға ие екенін көрсетті.
1961 жылы Крут пен Вайнберг өсірілген фибробласттарда in vitro галактоземияда фермент ақауын көрсетті.
1967 Зегмиллер және басқалар Леш-Нихан синдромында гипоксантин-гуанинфосфорибозилтрансфераза (HPRT) ақауын ашты.
1968 Кливер пигментті ксеродермадағы эксцизді жөндеудің бұзылуын сипаттады
1970 Нойфельд мукополисахаридтердің ыдырау жолдарын анықтауға мүмкіндік беретін мукополисахаридоздардың ферментативті ақауларын анықтады.
1974 Браун және Голдштейн отбасылық гиперхолестеринемияда гидроксиметилглутарилКоА-редуктазаның генетикалық анықталған шамадан тыс өндірілуі осы ферменттің (HMG) белсенділігін модуляциялайтын мембраналық локализацияланған төмен тығыздықты липопротеиндік рецепторлардың ақауына байланысты екенін дәлелдеді.
1977 Слай және басқалары манноза-6-фосфатты (лизосомалық ферменттердің құрамдас бөлігі ретінде) фибробласт рецепторларымен танитынын көрсетті. Генетикалық өңдеу ақауы лизосомалық ферменттердің байланысуын болдырмайды, нәтижесінде олардың цитоплазмаға бөлінуі және кейіннен плазмаға секрециясы бұзылады (I-жасушалық ауру)
12 4. Гендердің әрекеті
1980 Псевдохипопаратиреозда рецептор мен циклазаның қосылуын қамтамасыз ететін ақуыздың ақауы анықталды.
Әрбір тірі жасушада көптеген химиялық реакциялар жүреді. Ферменттер - ерекше және өте маңызды функциялары бар ақуыздар. Оларды биокатализаторлар деп атайды. Денедегі ең бастысы - биохимиялық реакцияларды жеделдету. Өзара әрекеттесуі осы молекулалармен катализденетін бастапқы реагенттер субстрат, ал соңғы қосылыстар өнімдер деп аталады.
Табиғатта фермент белоктары тек тірі жүйелерде жұмыс істейді. Бірақ қазіргі биотехнологияда клиникалық диагностика, фармацевтика және медицина, тазартылған ферменттер немесе олардың кешендері, сондай-ақ жүйенің жұмыс істеуі және зерттеуші үшін деректерді визуализациялау үшін қажетті қосымша компоненттер пайдаланылады.
Ферменттердің биологиялық маңызы және қасиеттері
Бұл молекулаларсыз тірі организм жұмыс істей алмайды. Барлық өмірлік процестер ферменттердің арқасында біркелкі жұмыс істейді. Ағзадағы ферменттік белоктардың негізгі қызметі – зат алмасуды реттеу. Оларсыз қалыпты метаболизм мүмкін емес. Молекулалардың белсенділігін реттеу активаторлардың (индукторлардың) немесе ингибиторлардың әсерінен жүреді. Басқару жұмыс істейді әртүрлі деңгейлерақуыз синтезі. Ол сондай-ақ дайын молекулада «жұмыс істейді».
Ферменттік ақуыздардың негізгі қасиеттері белгілі бір субстратқа спецификалық болып табылады. Және, тиісінше, тек бір немесе, сирек, реакциялар сериясын катализдеу мүмкіндігі. Әдетте мұндай процестер қайтымды. Бір фермент екі функцияға да жауап береді. Бірақ бұл бәрі емес.

Ферменттік ақуыздардың рөлі өте маңызды. Оларсыз биохимиялық реакциялар жүрмейді. Ферменттердің әсерінен реагенттер белсендіру кедергісін айтарлықтай энергия шығынынсыз жеңе алады. Дене температурасын 100 °C-тан жоғары қыздыру немесе агрессивті компоненттерді пайдалану мүмкін емес. химиялық зертхана. Фермент ақуызы субстратпен байланысады. Байланысты күйде модификация соңғысының кейіннен шығарылуымен жүреді. Химиялық синтезде қолданылатын барлық катализаторлар дәл осылай жұмыс істейді.
Белок-фермент молекуласының ұйымдасу деңгейлері қандай?
Әдетте бұл молекулалар үшіншілік (глобула) немесе төрттік (бірнеше байланысқан глобулалар) болады. белок құрылымы. Олар алдымен сызықты түрде синтезделеді. Содан кейін олар қажетті құрылымға құлайды. Белсенділікті қамтамасыз ету үшін биокатализаторға белгілі бір құрылым қажет.

Ферменттер, басқа белоктар сияқты, жылумен, рН шамадан тыс мәндерімен және агрессивті химиялық қосылыстармен жойылады.
Ферменттердің қосымша қасиеттері
Олардың ішінде компоненттердің келесі ерекшеліктері бар:
- Стереоспецификалық – бір ғана өнімнің қалыптасуы.
- Региоселективтілік – алшақтық химиялық байланыснемесе топты тек бір позицияда өзгерту.
- Хемоселективтілік – бір ғана реакцияның катализі.
Жұмыстың ерекшеліктері
Деңгейі өзгереді. Бірақ кез келген фермент белгілі бір субстратқа немесе құрылымы жағынан ұқсас қосылыстар тобына қарсы әрқашан белсенді. Ақуыз емес катализаторларда мұндай қасиет болмайды. Ерекшелік 10−10 моль/л жетуі мүмкін байланыс тұрақтысымен (моль/л) өлшенеді. Белсенді ферменттің жұмысы жылдам. Бір молекула секундына мыңдаған және миллиондаған операцияларды катализдейді. Биохимиялық реакциялардың үдеу дәрежесі кәдімгі катализаторларға қарағанда айтарлықтай (1000-100000 есе) жоғары.
Ферменттердің әрекеті бірнеше механизмдерге негізделген. Ең қарапайым әрекеттесу бір субстрат молекуласымен жүреді, содан кейін өнім түзіледі. Көптеген ферменттер реакцияға түсетін 2-3 түрлі молекуланы байланыстыруға қабілетті. Мысалы, топты немесе атомды бір қосылыстан екінші қосылысқа ауыстыру немесе «пинг-понг» принципі арқылы қосарлы алмастыру. Бұл реакцияларда әдетте бір субстрат біріктіріледі, ал екіншісі функционалды топ арқылы ферментке қосылады.
Зерттеу келесі әдістерді қолдана отырып жүргізіледі:
- Аралық және соңғы өнімдердің анықтамалары.
- Құрылымның геометриясын және субстратпен байланысты функционалдық топтарды зерттеу және жоғары қамтамасыз ету
- Фермент гендерінің мутациялары және оның синтезі мен белсенділігінің өзгеруін анықтау.

Белсенді және байланыстырушы сайт
Субстрат молекуласы мөлшері жағынан фермент ақуызына қарағанда әлдеқайда кіші. Сондықтан байланыс биокатализатордың функционалдық топтарының аз санына байланысты болады. Олар аминқышқылдарының белгілі бір жиынтығынан тұратын белсенді орталықты құрайды. Құрылымда протеинді емес табиғаттың протездік тобы бар, ол да белсенді орталықтың бөлігі болуы мүмкін.
Ферменттердің жеке тобын ажырату керек. Олардың молекуласында молекуламен үнемі байланысатын және одан бөлініп шығатын кофермент болады. Толық түзілген фермент ақуызын холофермент деп атайды, ал кофакторды алып тастағанда оны апофермент деп атайды. Витаминдер, металдар, азотты негіздердің туындылары (NAD – никотинамид адениндинуклеотиді, ФАД – флавин адениндинуклеотиді, ФМН – флавин мононуклеотиді) жиі кофермент ретінде әрекет етеді.

Байланыстыру орны субстраттың жақындық ерекшелігін қамтамасыз етеді. Оның арқасында тұрақты субстрат-ферменттік кешен түзіледі. Глобулдың құрылымы субстратты байланыстыруды қамтамасыз ете отырып, бетінде белгілі бір өлшемдегі тауашаны (жарық немесе ойық) болатындай етіп салынған. Бұл аймақ әдетте белсенді орталықтың жанында орналасады. Жеке ферменттерде кофакторлармен немесе металл иондарымен байланысу орындары бар.
Қорытынды
Ағзада ферменттік ақуыз маңызды рөл атқарады. Мұндай заттар химиялық реакцияларды катализдейді және метаболикалық процесс - метаболизмге жауап береді. Кез келген тірі жасушада жүздеген биохимиялық процестер үнемі жүреді, соның ішінде қалпына келтіру реакциялары, қосылыстардың бөлінуі және синтезі. Заттар энергияның көп бөлінуімен үнемі тотығады. Ол өз кезегінде көмірсулардың, белоктардың, майлардың және олардың кешендерінің түзілуіне жұмсалады. Ыдырау өнімдері қажетті органикалық қосылыстарды синтездеуге арналған құрылымдық элементтер болып табылады.
» , » Бір ген, бір фермент
Бір ген, бір фермент
92
Жарияланған күні: 2018 жылғы 24 шілде
Бір ген, бір фермент гипотезасы 1940 жылдардың басында алға қойылған идея, әрбір ген бір ферменттің синтезін немесе белсенділігін бақылайды. Генетика мен биохимияның салаларын біріктіретін концепцияны американдық генетик Джордж Уэллс Бидл мен американдық биохимигі Эдвард Л.Татум ұсынған, олар Neurospora crassa бойынша зерттеулер жүргізген. Олардың эксперименттері алдымен пішінді мутация тудыратын рентген сәулелеріне бейнелеуді, содан кейін оны жабайы типті штаммның өмір сүруіне қажетті маңызды қоректік заттардан тұратын минималды өсу ортасында өсіруді қамтыды. Олар мутантты зең штаммдарының өсуі үшін белгілі бір аминқышқылдарының қосылуын қажет ететінін анықтады. Осы ақпаратты пайдалана отырып, зерттеушілер белгілі бір гендердегі мутацияларды әдетте жетіспейтін аминқышқылдарын шығаратын метаболикалық жолдардағы арнайы ферменттердің бұзылуымен байланыстыра алды. Қазіргі уақытта барлық гендер ферментті кодтамайтыны және кейбір ферменттер екі немесе одан да көп гендермен кодталған бірнеше қысқа полипептидтерден тұратыны белгілі.
Эукариоттық гендердің экзон-интрондық ұйымының ашылуы және альтернативті сплайсинг мүмкіндігі біріншілік транскрипттің бірдей нуклеотидті тізбегі бірнеше полипептидтік тізбектердің синтезін қамтамасыз ете алатынын көрсетті. әртүрлі функцияларнемесе олардың өзгертілген аналогтары. Мысалы, ашытқы митохондрияларында респираторлық цитохром b ферментін кодтайтын қорап (немесе коб) гені бар. Ол екі түрде болуы мүмкін: 6400 бит-тен тұратын «ұзын» геннің жалпы ұзындығы 1155 бит болатын 6 экзон бар. және 5 интрон. Геннің қысқа түрі 3300 б.б. және 2 интроны бар. Бұл шын мәнінде алғашқы үш интрондары жоқ «ұзын» ген. Геннің екі түрі де бірдей жақсы экспрессияланған.
Алғашқы екі экзонның және екінші интронның нуклеотидтерінің бөлігінің біріктірілген нуклеотидтер тізбегіне негізделген «ұзын» қорап генінің бірінші интронын алып тастағаннан кейін тәуелсіз ақуыз - РНҚ матуразасы үшін матрица түзіледі (Cурет 1). 3.43). РНҚ матуразасының қызметі сплайсингтің келесі сатысын қамтамасыз ету болып табылады - біріншілік транскрипттен екінші интронды алып тастау және ең соңында b цитохромының шаблонын қалыптастыру.
Тағы бір мысал - лимфоциттердегі антидене молекулаларының құрылымын кодтайтын бастапқы транскрипттің сплайсинг үлгісінің өзгеруі. Антиденелердің мембраналық формасы С-соңында аминқышқылдарының ұзын «құйрығына» ие, ол мембранаға ақуыздың бекітілуін қамтамасыз етеді. Антиденелердің бөлінетін түрінде мұндай құйрық болмайды, бұл сплайсинг кезінде біріншілік транскрипттен осы аймақты кодтайтын нуклеотидтердің жойылуымен түсіндіріледі.
Вирустар мен бактерияларда бір ген бір уақытта басқа геннің бөлігі бола алатын немесе белгілі бір ДНҚ нуклеотидті тізбегі екі түрлі қабаттасатын гендердің бөлігі болуы мүмкін жағдай сипатталған. Мысалы, FX174 фагының геномының физикалық картасы (3.44-сурет) В генінің тізбегі А генінің ішінде орналасқанын, ал Е генінің D генінің тізбегінің бөлігі екенін көрсетеді. Фаг ұйымының бұл ерекшелігі. геном өзінің салыстырмалы түрде аз мөлшері (ол 5386 нуклеотидтен тұрады) мен барлық синтезделген белоктардағы аминқышқылдары қалдықтарының саны арасындағы бар сәйкессіздікті түсіндіре алды, бұл берілген геномдық сыйымдылық үшін теориялық тұрғыдан рұқсат етілгеннен асып түседі. Қабатталатын гендерден (A және B немесе E және D) синтезделген мРНҚ-да әртүрлі пептидтік тізбектерді жинақтау мүмкіндігі осы мРНҚ ішінде рибосомаларды байланыстыру орындарының болуымен қамтамасыз етіледі. Бұл басқа пептидті аударуды жаңа бастапқы нүктеден бастауға мүмкіндік береді.
В генінің нуклеотидтер тізбегі бір мезгілде А генінің бөлігі, ал Е гені D генінің бөлігі болып табылады.
Фреймді ауыстырумен де, бір оқу кадрында да аударылған қабаттасатын гендер λ фаг геномында да табылды. Сондай-ақ бір ДНҚ секциясының екі комплементарлы тізбегінен екі түрлі мРНҚ-ны транскрипциялауға болады деп болжанады. Бұл РНҚ-полимеразаның ДНҚ молекуласы бойымен әртүрлі бағытта қозғалуын анықтайтын промоторлық аймақтардың болуын талап етеді.
Бір ДНҚ тізбегінен әртүрлі ақпаратты оқудың рұқсат етілгендігін көрсететін сипатталған жағдайлар, гендердің қабаттасуы вирустардың және, мүмкін, прокариоттардың геномын ұйымдастырудың жеткілікті кең таралған элементі болып табылады. Эукариоттарда гендік үзіліс бір ДНҚ тізбегінен әртүрлі пептидтердің синтезіне де мүмкіндік береді.
Осының бәрін ескере отырып, геннің анықтамасына түзетулер енгізу қажет. Белгілі бір ақуызды бірегей кодтайтын ДНҚ-ның үздіксіз тізбегі ретіндегі ген туралы енді айта алмайтынымыз анық. Шамасы, қазіргі уақытта «Бір ген - бір полипептид» формуласы әлі де ең қолайлы болып саналуы керек, дегенмен кейбір авторлар оны өзгертуді ұсынады: «Бір полипептид - бір ген». Кез келген жағдайда ген терминін өзінің химиялық табиғаты бойынша полинуклеотид болып табылатын және полипептидтік тізбекті, тРНҚ немесе рРНҚ синтездеу мүмкіндігін анықтайтын тұқым қуалайтын материалдың функционалдық бірлігі ретінде түсіну керек.
Бір ген, бір фермент.
1940 жылы Дж.Бидл мен Эдвард Татум гендердің метаболизмді қалай қамтамасыз ететінін зерттеу үшін жаңа тәсілді неғұрлым ыңғайлы зерттеу нысанында – микроскопиялық Neurospora crassa саңырауқұлақтарында қолданды.Олар мутацияларды алды, оларда; сол немесе басқа метаболикалық ферменттің белсенділігі байқалмады. Ал бұл мутантты саңырауқұлақтың белгілі бір метаболитті өздігінен синтездей алмайтындығына әкелді (мысалы, лейцин амин қышқылы) және қоректік ортаға лейцинді қосқанда ғана өмір сүре алады. Дж.Бидл мен Э.Татум тұжырымдаған «бір ген, бір фермент» теориясы тез арада генетиктер арасында кеңінен танылып, олардың өздері Нобель сыйлығына ие болды.
Әдістері. Әртүрлі метаболикалық жолдарды қамтамасыз ететін ферменттердің әрекетінің бұзылуына әкелетін «биохимиялық мутациялар» деп аталатындарды таңдау ғылым үшін ғана емес, сонымен қатар тәжірибе үшін де өте жемісті болды. Біріншіден, олар өнеркәсіптік микроорганизмдердің генетикасы мен селекциясының пайда болуына, содан кейін антибиотиктер, витаминдер, аминқышқылдары және т.б. сияқты стратегиялық маңызды заттарды шамадан тыс өндіретін микроорганизмдердің штаммдарын қолданатын микробиологиялық өнеркәсіпке әкелді.Селекция және гендік инженерия принциптері. суперпродуценттердің штаммдары «бір ген бір ферментті кодтайды» деген идеяға негізделген. Бұл идея тәжірибе үшін тамаша болса да, миллиондаған доллар пайда әкеледі және миллиондаған адамдардың өмірін (антибиотиктер) сақтайды - бұл түпкілікті емес. Бір ген бір ғана фермент емес.
| " |
Генетика- ғылым жас емес, ондағы зерттеулер 1865 жылы Мендельден бастап бүгінгі күнге дейін бірнеше ғасырлар бойы жүргізілді. Тұқым қуалайтын сипаттама бірлігіне арналған «ген» терминін алғаш рет 1911 жылы Иогансен ұсынған, ал 1940 жылдары Татум мен Бидл ұсынған «бір ген, бір фермент» тұжырымдамасы арқылы нақтыланған.
Бұл позиция Дрозофила шыбындарына жүргізілген эксперименттерде анықталды, бірақ адамдарға бірдей қатысты; Сайып келгенде, барлық тіршілік иелерінің өмірі олардың ДНҚ-сы арқылы анықталады. Адамның ДНҚ молекуласы барлық басқа организмдерге қарағанда үлкенірек, құрылысы күрделірек, бірақ оның қызметтерінің мәні барлық тірі организмдерде бірдей.
тұжырымдамасы» бір ген – бір ферментТатум мен Бидлдің идеялары негізінде пайда болған ", келесідей тұжырымдауға болады:
1. Барлығы биологиялық процестергенетикалық бақылауда болады.
2. Барлық биохимиялық процестер сатылы реакциялар түрінде жүреді.
3. Әрбір биохимиялық реакция түптеп келгенде әртүрлі жеке гендердің бақылауында болады.
4. Белгілі бір гендегі мутация жасушаның белгілі бір химиялық реакцияны жүргізу қабілетінің өзгеруіне әкеледі.
Содан бері «бір ген – бір фермент» ұғымы біршама кеңейіп, қазір « бір ген – бір белок" Сонымен қатар, жақында жүргізілген зерттеулер кейбір гендер бірегей ақуыздарды өндіру үшін басқалармен бірге әрекет ететінін көрсетеді, яғни кейбір гендер бірнеше ақуызды кодтай алады.
Адам геномышамамен 3 миллиард жұп нуклеотидтерден тұрады; онда 50 000-нан 100 000-ға дейін бар деп есептеледі. Геномды ашқаннан кейін бар болғаны 30 000-ға жуық ген бар екені белгілі болды.Бұл гендердің өзара әрекеттесуі күтілгеннен әлдеқайда күрделі. Гендер ДНҚ тізбегінде кодталады, олар белгілі бір ядролық ақуыздармен қосылып хромосома түзеді.
Гендер- жай ғана ДНҚ сегменттері емес: олар кодтаушы тізбектер - экзондар арқылы түзіледі, кодталмаған тізбектермен - нитрондармен қиылысады. Экзондар ДНҚ-ның экспрессиялық бөлігі ретінде организмнің ең маңызды молекуласының аз ғана бөлігін құрайды; оның көп бөлігі экспрессияланбайды, нитрондардан түзіледі және көбінесе «үнсіз» ДНҚ деп аталады.
Шамамен мөлшері мен құрылымы адам геномытөмендегі суретте берілген. Адам хромосомасының функционалдық ұзындығы центиморганидтермен өрнектеледі. Центиморганид (cM) - мейоз кезінде қиылысу ықтималдығы 1% болатын қашықтық. Гендердің байланысын талдау адам геномының ұзындығы шамамен 3000 см болатынын көрсетті.
Орташа хромосома 130 миллион нуклеотидтік негіз жұбында кодталған шамамен 1500 генді қамтиды. Төмендегі суретте геномның физикалық және функционалдық өлшемдері схемалық түрде көрсетілген: біріншісі нуклеотидтік жұптарда, ал екіншісі cM-де есептеледі. Адам геномының көп бөлігі «үнсіз» ДНҚ-мен ұсынылған және экспрессияланбайды.
Қосулы ДНҚ матрицасыТранскрипция процесінің нәтижесінде РНҚ, содан кейін белок синтезделеді. Демек, ДНҚ тізбегі жасушаның функционалды белоктарының ретін толығымен анықтайды. Барлық белоктар келесідей синтезделеді:
ДНҚ => РНҚ => ақуыз

Адамдардың және басқа сүтқоректілердің генетикалық аппараты басқа тірі организмдерге қарағанда күрделірек, өйткені сүтқоректілердің кейбір гендерінің бөлімдері басқаларының бөліктерімен біріктірілуі мүмкін. гендер, нәтижесінде мүлдем жаңа ақуыз синтезі немесе бөлек жасушалық функцияны басқару.
Демек, адамдарда экспрессияланған гендердің көлемін іс жүзінде арттырмай-ақ көбейтуге болады. ДНҚнемесе гендердің абсолютті саны.
Тұтастай алғанда, барлық генетикалық материалдың шамамен 70% -ы экспрессияланбайды.
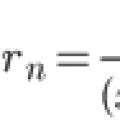 Тейлор қатарын кеңейту. Кәдімгі үшін Коши есебінің шамамен шешімі
Тейлор қатарын кеңейту. Кәдімгі үшін Коши есебінің шамамен шешімі Мектепте не ОҚЫТЫЛМАЙДЫ Мектепте нені оқытпайды деп сұраңыз
Мектепте не ОҚЫТЫЛМАЙДЫ Мектепте нені оқытпайды деп сұраңыз Ақшаны ойлау формуласы (А
Ақшаны ойлау формуласы (А