یک ژن یک آنزیم تفسیر مدرن. تئوری یک ژن - یک آنزیم
4.2.1. فرضیه "یک ژن، یک آنزیم".
تحقیق اولپس از اینکه گارود در سال 1902 اشاره کرد که نقص ژنتیکی در آلکاپتونوری با ناتوانی بدن در تجزیه اسید هموژنتیزیک مرتبط است، روشن کردن مکانیسم خاص این اختلال مهم بود. از آنجایی که در آن زمان شناخته شده بود که واکنش های متابولیک توسط آنزیم ها کاتالیز می شود، می توان فرض کرد که اختلال در برخی از آنزیم ها منجر به آلکاپتونوری می شود. چنین فرضیه ای توسط دریش (در سال 1896) مورد بحث قرار گرفت. همچنین توسط Haldane (1920، نگاه کنید) و Garrod (1923) بیان شد. مراحل مهمتوسعه ژنتیک بیوشیمیایی با کار کوچن و بوتناندت بر روی مطالعه رنگ چشم در پروانه آسیاب آغاز شد. افستیا کوهنیلاو مطالعات مشابه توسط Beadle و Ephrussi در مورد مگس سرکه(1936). در این آثار پیشگام، جهشیافتههای حشرهای که قبلاً با روشهای ژنتیکی مورد مطالعه قرار گرفته بودند، برای روشن کردن مکانیسمهای عمل ژن انتخاب شدند. با این حال، این رویکرد به موفقیت منجر نشد. معلوم شد که مشکل خیلی پیچیده است و برای حل آن لازم بود:
1) یک ارگانیسم مدل ساده مناسب برای مطالعه تجربی را انتخاب کنید.
2) به دنبال اساس ژنتیکی صفات بیوشیمیایی، به جای مبنای بیوشیمیایی صفات تعیین شده ژنتیکی. هر دو شرط در کار Beadle و Tatum در سال 1941 برآورده شد (همچنین به Beadle، 1945 مراجعه کنید).
مدل Beadle و Tatum. مقاله این محققین اینگونه آغاز شد:
از دیدگاه ژنتیک فیزیولوژیکی، رشد و عملکرد یک ارگانیسم را می توان به سیستم پیچیده ای از واکنش های شیمیایی که به نوعی توسط ژن ها کنترل می شوند، تقلیل داد. کاملاً منطقی است که فرض کنیم این ژن ها یا خود به عنوان آنزیم عمل می کنند یا ویژگی آنها را تعیین می کنند. مشخص است که ژنتیک-فیزیولوژیست ها معمولاً سعی می کنند اساس فیزیولوژیکی و بیوشیمیایی صفات ارثی از قبل شناخته شده را بررسی کنند. این رویکرد امکان ایجاد این تعداد بیو را فراهم کرد واکنش های شیمیاییتوسط ژن های خاص کنترل می شود. چنین مطالعاتی نشان داده است که آنزیم ها و ژن ها دارای ویژگی یکسانی هستند. با این حال، قابلیت های این رویکرد محدود است. جدی ترین محدودیت این است که در این مورد، محققان به صفات ارثی توجه می کنند که اثر کشنده ای ندارند و بنابراین با واکنش هایی همراه هستند که برای زندگی ارگانیسم چندان مهم نیستند. مشکل دوم این است که رویکرد سنتی به مشکل شامل استفاده از نشانه های ظاهری بیرونی است. بسیاری از آنها تغییرات مورفولوژیکی مبتنی بر سیستمهای واکنش بیوشیمیایی آنقدر پیچیده هستند که تجزیه و تحلیل آنها به طور غیرعادی دشوار است.
ملاحظات مشابه ما را به نتیجه گیری زیر هدایت کرد. مطالعه مشکل کلی کنترل ژنتیکی واکنشهای بیوشیمیایی که توسعه و متابولیسم را تعیین میکنند باید با استفاده از آن انجام شود رویه برخلاف روش پذیرفته شده عمومی:به جای تلاش برای یافتن اساس شیمیایی صفات ارثی شناخته شده، لازم است که مشخص شود آیا و چگونه ژن ها واکنش های بیوشیمیایی شناخته شده را کنترل می کنند.نوروسپورا که متعلق به آسکومیست ها است دارای خواصی است که اجرای چنین رویکردی را ممکن می کند و در عین حال به عنوان یک شی مناسب برای تحقیقات ژنتیکی عمل می کند. به همین دلیل است که برنامه ما بر اساس استفاده از این ارگانیسم خاص ساخته شده است. ما فرض کردیم که قرار گرفتن در معرض اشعه ایکس باعث جهش در ژن هایی می شود که واکنش های شیمیایی خاصی را کنترل می کنند. فرض کنید برای زنده ماندن در یک محیط معین، یک موجود زنده باید نوعی واکنش شیمیایی را انجام دهد، در این صورت یک جهش یافته فاقد چنین توانایی در این شرایط غیرقابل زنده ماندن خواهد بود. با این حال، اگر در محیطی رشد کند که محصول حیاتی یک واکنش ژنتیکی مسدود شده به آن اضافه شده باشد، قابل نگهداری و مطالعه است.
4 عمل ژن ها 9
برنج. 4.1. طرح آزمایشی برای شناسایی جهش یافته های بیوشیمیایی نوروسپورا در یک محیط کامل، جهش های ناشی از اشعه ایکس یا اشعه ماوراء بنفش رشد قارچ را مختل نمی کند. با این حال، جهش یافته در حداقل محیط رشد نمی کند. هنگامی که ویتامین ها به یک محیط حداقل افزوده می شود، توانایی رشد بازیابی می شود. وقتی اسیدهای آمینه اضافه می شوند، رشدی وجود ندارد. بر اساس این داده ها، می توان فرض کرد که جهش در ژنی رخ داده است که متابولیسم ویتامین را کنترل می کند. گام بعدی شناسایی ویتامینی است که می تواند عملکرد طبیعی خود را بازگرداند.یک بلوک ژنتیکی در میان واکنش های بیوسنتز ویتامین یافت می شود.سپس، Beadle و Tatum طرح آزمایشی را توصیف می کنند (شکل 4.1). محیط کامل حاوی آگار، نمک های معدنی، عصاره مالت، عصاره مخمر و گلوکز بود. حداقل محیط فقط حاوی آگار، نمک، بیوتین و منبع کربن بود. جهش یافته هایی که در محیط کامل رشد کردند و در محیط حداقل رشد نکردند با جزئیات بیشتر مورد مطالعه قرار گرفتند. برای شناسایی ترکیبی که سنتز آن در هر یک از جهشیافتهها مختل شده بود، اجزای جداگانه محیط کامل به حداقل آگار اضافه شد.
به این ترتیب، سویه هایی جدا شدند که قادر به سنتز فاکتورهای رشد خاصی نبودند: پیریدوکسین، تیامین و پاراآمینوبنزوئیک اسید. نشان داده شده است که این نقایص ناشی از جهش در مکان های خاص است. این کار پایه و اساس مطالعات متعددی را بر روی نوروسپورا، باکتری ها و مخمرها ایجاد کرد که در آنها مطابقت "بلوک های ژنتیکی" مسئول مراحل متابولیک فردی و اختلالات آنزیمی خاص ایجاد شد. این رویکرد به سرعت به ابزاری تبدیل شده است که به محققان امکان می دهد مسیرهای متابولیک را کشف کنند.
فرضیه "یک ژن - یک آنزیم" تایید تجربی قوی دریافت کرده است. همانطور که کار دهه های بعدی نشان داد، به طرز شگفت انگیزی پربار بود. تجزیه و تحلیل آنزیم های معیوب و انواع طبیعی آنها به زودی امکان شناسایی دسته ای از اختلالات ژنتیکی را فراهم کرد که منجر به تغییراتی در عملکرد آنزیم شد، اگرچه خود پروتئین هنوز قابل تشخیص بود و خواص ایمنی خود را حفظ کرد. در موارد دیگر، دمای بهینه برای فعالیت آنزیم تغییر کرد. برخی از انواع را می توان با یک جهش توضیح داد که بر یک مکانیسم تنظیمی کلی تأثیر می گذارد و در نتیجه فعالیت کل گروهی از آنزیم ها را تغییر می دهد. مطالعات مشابه منجر به ایجاد مفهوم تنظیم فعالیت ژن در باکتری ها شد که شامل مفهوم اپرون بود.
10 4. عمل ژن ها
اولین نمونه از اختلالات آنزیمی در انسان.اولین بیماری ارثی انسانی که اختلال آنزیمی برای آن نشان داده شد، متهموگلوبینمی با حالت ارثی مغلوب بود (گیبسون و هریسون، 1947؛ گیبسون، 1948) (25080). در این مورد، آنزیم آسیب دیده متهموگلوبین ردوکتاز وابسته به NADH است. اولین تلاش برای مطالعه سیستماتیک گروهی از بیماری های انسانی مرتبط با نقص متابولیک در سال 1951 انجام شد. در مطالعه ای در مورد بیماری ذخیره گلیکوژن، Coreys نشان داد که در هشت مورد از ده مورد وضعیت پاتولوژیک که به عنوان بیماری گیرک تشخیص داده شد (23220)، ساختار گلیکوژن کبد طبیعی بود و در دو مورد به وضوح غیر طبیعی بود. همچنین واضح بود که گلیکوژن کبد که بیش از حد تجمع می یابد، نمی تواند مستقیماً به قند تبدیل شود، زیرا بیماران به هیپوگلیسمی تمایل داشتند. آنزیم های زیادی برای تجزیه گلیکوژن برای تشکیل گلوکز در کبد مورد نیاز است. دو مورد از آنها، آمیلو-1،6-گلوکوزیداز و گلوکز 6-فسفاتاز، برای مطالعه به عنوان عناصر معیوب احتمالی سیستم آنزیمی انتخاب شدند. آزادسازی فسفات از گلوکز-6 فسفات در هموژنه های کبد در مقادیر مختلف pH اندازه گیری شد. نتایج در شکل ارائه شده است. 4.2. در کبد نرمال، فعالیت بالا با بهینه در pH 6-7 یافت شد. اختلال عملکرد شدید کبد در سیروز با کاهش جزئی فعالیت مرتبط است. از سوی دیگر، در مورد بیماری گیرک با پیامد کشنده، فعالیت آنزیم به هیچ وجه قابل تشخیص نیست. همین نتیجه هنگام معاینه یک بیمار مشابه دوم به دست آمد. دو بیمار با علائم کمتر شدید کاهش قابل توجهی در فعالیت داشتند.
نتیجه گیری شد که در این موارد از بیماری کشنده گیرک نقص گلوکز-6-فسفاتاز وجود دارد. با این حال، در بیشتر موارد خفیف تر، فعالیت این آنزیم کمتر از سیروز کبدی نبود و تنها در دو بیمار کمی کمتر بود (شکل 4.2).
به گفته Coreys، تجمع غیرطبیعی گلیکوژن در بافت عضلانی نمی تواند با کمبود گلوکز-6-فسفاتاز مرتبط باشد، زیرا این آنزیم در ماهیچه ها وجود ندارد و طبیعی است. آنها اختلال در فعالیت آمیلو-1،6-گلوکوزیداز را به عنوان توضیحی ممکن برای گلیکوژنوز عضلانی پیشنهاد کردند. این پیشبینی به زودی تأیید شد: فوربس چنین نقصی را در یکی از موارد بالینی مهم بیماری ذخیرهسازی گلیکوژن که عضلات قلبی و اسکلتی را درگیر میکند، کشف کرد. حالا ما
4. عمل ژن ها 11
تعداد زیادی نقص آنزیمی در بیماری ذخیره گلیکوژن شناخته شده است.
اگرچه اشکال مختلف این بیماری تا حدودی در درجه تظاهرات متفاوت است، اما از نظر بالینی مشترکات زیادی دارند. با یک استثنا، همه آنها به روش اتوزومال مغلوب به ارث می رسند. اگر نقایص آنزیمی کشف نشده بود، آسیب شناسی ذخیره گلیکوژن به عنوان یکی از بیماری ها با همبستگی های درون خانواده مشخصه در شدت، جزئیات علائم و زمان مرگ در نظر گرفته می شود. بنابراین، ما نمونهای داریم که در آن ناهمگنی ژنتیکی، که فقط بر اساس مطالعه فنوتیپ (بخش 3.3.5) قابل فرض است، با تجزیه و تحلیل در سطح بیوشیمیایی تأیید شد: تحقیق. فعالیت آنزیمیامکان شناسایی ژن های خاص را فراهم کرد.
در سال های بعد، سرعت تحقیقات در زمینه نقص های آنزیمی افزایش یافت و برای 588 ژن اتوزومال مغلوب شناسایی شده که مک کوزیک در ویرایش ششم کتاب خود وراثت مندلی در انسان (1983) توضیح می دهد، نقص های آنزیمی خاصی در بیش از 170 مورد پیشرفت ما در این زمینه ارتباط مستقیمی با توسعه مفاهیم و روش های ژنتیک مولکولی دارد.
برخی از مراحل مطالعه اختلالات آنزیمی در انسان.ما فقط مهم ترین نقاط عطف این فرآیند در حال انجام را ارائه می کنیم: 1934 فولینگ فنیل کتونوری را کشف کرد.
1941 Beadle و Tatum فرضیه "یک ژن، یک آنزیم" را فرموله کردند 1948 Gibson اولین مورد اختلال آنزیمی در یک بیماری انسانی (متهموگلوبینمی مغلوب) را تشریح کرد.
1952 Coreys کمبود گلوکز-6-فسفاتاز را در بیماری گیرک کشف کرد.
1953 جرویس عدم وجود فنیل آلانین هیدروکسیلاز را در فنیل کتونوری نشان داد. بیکل اولین تلاش را برای کاهش یک اختلال آنزیمی با استفاده از رژیم غذایی کم فنیل آلانین گزارش کرد
1955 Smithies تکنیک الکتروفورز ژل نشاسته را توسعه داد
1956 کارسون و همکاران نقصی در گلوکز-6-فسفات دهیدروژناز (G6PD) در مورد کم خونی همولیتیک القایی کشف کردند.
1957 Kalkar و همکاران کمبود آنزیمی را در گالاکتوزمیا توصیف کردند و نشان دادند که انسان و باکتری دارای نقصهای یکسانی در فعالیت آنزیمی هستند.
1961 کروت و واینبرگ نقص آنزیمی را در گالاکتوزمی در شرایط آزمایشگاهی در فیبروبلاست های کشت شده نشان دادند.
1967 Seegmiller و همکاران یک نقص هیپوگزانتین-گوانین فسفریبوزیل ترانسفراز (HPRT) را در سندرم Lesch-Nyhan کشف کردند.
Cleaver در سال 1968 اختلالی در ترمیم اکسیزیون در خشکی پوستی پیگمنتوزوم را توصیف کرد
1970 نوفلد نقص آنزیمی را در موکوپلی ساکاریدوزها شناسایی کرد که امکان شناسایی مسیرهای تجزیه موکوپلی ساکاریدها را فراهم کرد.
1974 براون و گلدشتاین ثابت کردند که تولید بیش از حد تعیین شده ژنتیکی هیدروکسی متیل گلوتاریل کوآ ردوکتاز در هیپرکلسترولمی خانوادگی به دلیل نقص در گیرنده لیپوپروتئین با چگالی کم غشایی است که فعالیت این آنزیم (HMG) را تعدیل می کند.
1977 Sly و همکاران نشان دادند که مانوز-6-فسفات (به عنوان جزئی از آنزیم های لیزوزومی) توسط گیرنده های فیبروبلاست شناسایی می شود. نقص پردازش ژنتیکی از اتصال آنزیم های لیزوزومی جلوگیری می کند، در نتیجه آزاد شدن آنها در سیتوپلاسم و ترشح بعدی در پلاسما مختل می شود (بیماری سلول I).
12 4. عمل ژن ها
1980 در شبه هیپوپاراتیروئیدیسم، نقصی در پروتئینی که جفت شدن گیرنده و سیکلاز را تضمین می کند، کشف شد.
بسیاری از واکنش های شیمیایی در هر سلول زنده رخ می دهد. آنزیم ها پروتئین هایی هستند که عملکردهای ویژه و بسیار مهمی دارند. به آنها بیوکاتالیست می گویند. نکته اصلی در بدن تسریع واکنش های بیوشیمیایی است. معرفهای اولیه که برهمکنش آنها توسط این مولکولها کاتالیز می شود، سوبسترا و ترکیبات نهایی محصول نامیده می شوند.
در طبیعت، پروتئین های آنزیمی فقط در سیستم های زنده کار می کنند. اما در بیوتکنولوژی مدرن، تشخیص بالینی، داروها و داروها، آنزیم های خالص شده یا کمپلکس های آنها و همچنین اجزای اضافی لازم برای عملکرد سیستم و تجسم داده ها برای محقق استفاده می شود.
اهمیت بیولوژیکی و خواص آنزیم ها
بدون این مولکول ها، یک موجود زنده قادر به فعالیت نخواهد بود. همه فرآیندهای حیاتی به لطف آنزیم ها به آرامی کار می کنند. وظیفه اصلی پروتئین های آنزیمی در بدن تنظیم متابولیسم است. بدون آنها متابولیسم طبیعی غیرممکن است. تنظیم فعالیت مولکول ها تحت تأثیر فعال کننده ها (القاء کننده ها) یا بازدارنده ها اتفاق می افتد. کنترل فعال است سطوح مختلفسنتز پروتئین. همچنین روی یک مولکول آماده "کار می کند".
خواص اصلی پروتئین های آنزیمی اختصاصی بودن به یک بستر خاص است. و بر این اساس، توانایی کاتالیز کردن تنها یک یا، کمتر، یک سری واکنش. معمولا چنین فرآیندهایی برگشت پذیر هستند. یک آنزیم مسئول هر دو عملکرد است. اما این همه ماجرا نیست.

نقش پروتئین های آنزیمی ضروری است. بدون آنها، واکنش های بیوشیمیایی رخ نمی دهد. با توجه به عمل آنزیم ها، این امکان برای معرف ها فراهم می شود که بدون صرف انرژی قابل توجه بر سد فعال سازی غلبه کنند. بدن نمی تواند دمای بالای 100 درجه سانتیگراد را گرم کند یا از اجزای تهاجمی مانند استفاده کند. آزمایشگاه شیمی. پروتئین آنزیمی به سوبسترا متصل می شود. در حالت محدود، اصلاح با انتشار بعدی دومی رخ می دهد. همه کاتالیزورهای مورد استفاده در سنتز شیمیایی دقیقاً اینگونه عمل می کنند.
سطوح سازماندهی یک مولکول پروتئین-آنزیم چیست؟
به طور معمول این مولکول ها دارای یک گلبول سوم (گلبول) یا چهارتایی (چند گلبول متصل) هستند. ساختار پروتئین. آنها ابتدا به صورت خطی سنتز می شوند. و سپس در ساختار مورد نیاز فرو می ریزند. برای اطمینان از فعالیت، یک بیوکاتالیست به ساختار خاصی نیاز دارد.

آنزیم ها مانند سایر پروتئین ها در اثر گرما، مقادیر زیاد pH و ترکیبات شیمیایی تهاجمی از بین می روند.
خواص اضافی آنزیم ها
از جمله ویژگی های اجزای زیر است:
- Stereospecificity - تشکیل تنها یک محصول.
- انتخاب منطقه - شکاف پیوند شیمیایییا اصلاح گروه فقط در یک موقعیت.
- انتخاب شیمیایی - کاتالیز تنها یک واکنش.
ویژگی های کار
سطح متفاوت است. اما هر آنزیمی همیشه در برابر یک سوبسترا یا گروهی از ترکیبات مشابه فعال است. کاتالیزورهای غیر پروتئینی این خاصیت را ندارند. ویژگی با ثابت اتصال (mol/L) اندازه گیری می شود که می تواند به 10-10 mol/L برسد. کار آنزیم فعال سریع است. یک مولکول هزاران تا میلیون ها عملیات را در ثانیه کاتالیز می کند. درجه شتاب واکنش های بیوشیمیایی به طور قابل توجهی (1000-100000 برابر) بیشتر از کاتالیزورهای معمولی است.
عمل آنزیم ها بر اساس مکانیسم های مختلفی است. ساده ترین برهمکنش با یک مولکول بستر و به دنبال آن تشکیل یک محصول اتفاق می افتد. اکثر آنزیم ها قادر به اتصال 2-3 مولکول مختلف هستند که وارد واکنش می شوند. به عنوان مثال، انتقال یک گروه یا اتم از یک ترکیب به ترکیب دیگر یا جایگزینی مضاعف با استفاده از اصل "پینگ پنگ". در این واکنش ها، یک سوبسترا معمولاً ترکیب می شود و دومی از طریق یک گروه عاملی به آنزیم متصل می شود.
مطالعه با استفاده از روش های زیر انجام می شود:
- تعاریف محصولات میانی و نهایی
- بررسی هندسه سازه و گروه های عملکردی مرتبط با بستر و تامین بالا
- جهش ژن های آنزیم و تعیین تغییرات در سنتز و فعالیت آن.

سایت فعال و الزام آور
مولکول سوبسترا از نظر اندازه بسیار کوچکتر از پروتئین آنزیم است. بنابراین، اتصال به دلیل تعداد کمی از گروه های عملکردی بیوکاتالیست رخ می دهد. آنها یک مرکز فعال متشکل از مجموعه خاصی از اسیدهای آمینه را تشکیل می دهند. این ساختار شامل یک گروه مصنوعی از طبیعت غیر پروتئینی است که می تواند بخشی از مرکز فعال نیز باشد.
گروه جداگانه ای از آنزیم ها باید متمایز شوند. مولکول آنها حاوی کوآنزیمی است که دائماً به مولکول متصل می شود و از آن آزاد می شود. پروتئین آنزیمی کاملاً تشکیل شده را هولوآنزیم می نامند و هنگامی که کوفاکتور حذف شود، آپوآنزیم نامیده می شود. ویتامین ها، فلزات، مشتقات بازهای نیتروژن دار (NAD - نیکوتین آمید آدنین دی نوکلئوتید، FAD - فلاوین آدنین دی نوکلئوتید، FMN - فلاوین مونوکلئوتید) اغلب به عنوان کوآنزیم عمل می کنند.

محل اتصال ویژگی میل ترکیبی بستر را فراهم می کند. با توجه به آن، یک کمپلکس سوبسترا-آنزیم پایدار تشکیل می شود. ساختار گلبول به گونه ای ساخته شده است که دارای یک طاقچه (شکاف یا فرورفتگی) با اندازه معینی بر روی سطح باشد و از اتصال زیرلایه اطمینان حاصل کند. این ناحیه معمولاً در نزدیکی مرکز فعال قرار دارد. آنزیم های منفرد دارای مکان هایی برای اتصال به کوفاکتورها یا یون های فلزی هستند.
نتیجه
پروتئین آنزیمی نقش مهمی در بدن دارد. چنین موادی واکنش های شیمیایی را کاتالیز می کنند و مسئول فرآیند متابولیک - متابولیسم هستند. در هر سلول زنده، صدها فرآیند بیوشیمیایی به طور مداوم رخ می دهد، از جمله واکنش های کاهشی، برش و سنتز ترکیبات. مواد به طور مداوم با آزاد شدن انرژی زیادی اکسید می شوند. این به نوبه خود برای تشکیل کربوهیدرات ها، پروتئین ها، چربی ها و مجتمع های آنها صرف می شود. محصولات تجزیه عناصر ساختاری برای سنتز ترکیبات آلی ضروری هستند.
»، » یک ژن، یک آنزیم
یک ژن، یک آنزیم
92
تاریخ انتشار: 24 جولای 2018
فرضیه یک ژن، یک آنزیم این ایده است که در اوایل دهه 1940 مطرح شد که هر ژن سنتز یا فعالیت یک آنزیم را کنترل می کند. این مفهوم که ترکیبی از زمینههای ژنتیک و بیوشیمی است، توسط جورج ولز بیدل، ژنتیکدان آمریکایی و ادوارد ال. تاتوم، بیوشیمیدان آمریکایی، که تحقیقاتی در مورد Neurospora crassa انجام دادند، پیشنهاد شد. آزمایشهای آنها ابتدا شامل تصویربرداری از فرم با اشعه ایکس القاکننده جهش و سپس کشت آن در محیط رشد حداقلی بود که فقط حاوی مواد مغذی ضروری برای بقای سویه نوع وحشی بود. آنها کشف کردند که گونه های کپک جهش یافته برای رشد نیاز به افزودن اسیدهای آمینه خاصی دارند. با استفاده از این اطلاعات، محققان توانستند جهش در ژنهای خاص را به اختلال در آنزیمهای خاص در مسیرهای متابولیکی که معمولاً اسیدهای آمینه از دست رفته را تولید میکنند، مرتبط کنند. اکنون مشخص شده است که همه ژن ها یک آنزیم را رمزگذاری نمی کنند و برخی از آنزیم ها از چندین پلی پپتید کوتاه تشکیل شده اند که توسط دو یا چند ژن رمزگذاری شده اند.
اکتشافات سازمان دهی اگزون-اینترون ژن های یوکاریوتی و امکان پیرایش جایگزین نشان داد که همان توالی نوکلئوتیدی رونوشت اولیه می تواند سنتز چندین زنجیره پلی پپتیدی را تضمین کند. توابع مختلفیا آنالوگ های اصلاح شده آنها. به عنوان مثال، میتوکندری مخمر حاوی یک ژن جعبه (یا بلال) است که آنزیم تنفسی سیتوکروم b را کد می کند. این می تواند به دو شکل وجود داشته باشد: ژن طولانی، متشکل از 6400 جفت باز، دارای 6 اگزون با طول کل 1155 جفت باز است. و 5 اینترون شکل کوتاه ژن شامل 3300 جفت باز است. و دارای 2 اینترون است. این در واقع یک ژن "طولانی" است که فاقد سه اینترون اول است. هر دو شکل ژن به یک اندازه به خوبی بیان می شوند.
پس از حذف اولین اینترون از ژن جعبه "طولانی"، بر اساس توالی نوکلئوتیدی ترکیبی دو اگزون اول و بخشی از نوکلئوتیدهای اینترون دوم، ماتریکسی برای پروتئین مستقل - RNA ماتاز تشکیل می شود (شکل 2). 3.43). عملکرد RNA maturase اطمینان از مرحله بعدی پیرایش - حذف اینترون دوم از رونوشت اولیه و در نهایت تشکیل یک الگو برای سیتوکروم b است.
مثال دیگر تغییر در الگوی پیرایش رونوشت اولیه است که ساختار مولکول های آنتی بادی در لنفوسیت ها را کد می کند. شکل غشایی آنتی بادی ها دارای یک "دم" بلند از اسیدهای آمینه در انتهای C است که تثبیت پروتئین روی غشاء را تضمین می کند. شکل ترشح شده آنتی بادی ها چنین دنباله ای ندارد، که با حذف نوکلئوتیدهای کد کننده این ناحیه از رونوشت اولیه در حین اتصال توضیح داده می شود.
در ویروس ها و باکتری ها، وضعیتی توصیف شده است که در آن یک ژن می تواند همزمان بخشی از یک ژن دیگر باشد، یا یک توالی نوکلئوتیدی DNA خاص می تواند بخشی از دو ژن مختلف همپوشانی باشد. به عنوان مثال، نقشه فیزیکی ژنوم فاژ FX174 (شکل 3.44) نشان می دهد که توالی ژن B در داخل ژن A قرار دارد و ژن E بخشی از توالی ژن D است. این ویژگی سازمان فاژ است. ژنوم توانست اختلاف موجود بین اندازه نسبتا کوچک خود (شامل 5386 نوکلئوتید) و تعداد باقیماندههای اسید آمینه در تمام پروتئینهای سنتز شده را توضیح دهد، که بیش از حد مجاز برای ظرفیت ژنوم معین است. امکان مونتاژ زنجیرههای پپتیدی مختلف بر روی mRNA سنتز شده از ژنهای همپوشانی (A و B یا E و D) با حضور مکانهای اتصال ریبوزوم در این mRNA تضمین میشود. این اجازه می دهد تا ترجمه پپتید دیگر از یک نقطه شروع جدید شروع شود.
توالی نوکلئوتیدی ژن B به طور همزمان بخشی از ژن A و ژن E بخشی از ژن D است.
ژنهای همپوشانی، که هم با تغییر چارچوب و هم در یک چارچوب خواندن ترجمه شدهاند، در ژنوم فاژ λ نیز یافت شدند. همچنین فرض بر این است که امکان رونویسی دو mRNA مختلف از هر دو رشته مکمل یک بخش DNA وجود دارد. این امر مستلزم وجود مناطق پروموتر است که حرکت RNA پلیمراز را در جهات مختلف در طول مولکول DNA تعیین می کند.
موقعیت های توصیف شده، که نشان دهنده مجاز بودن خواندن اطلاعات مختلف از یک توالی DNA است، نشان می دهد که ژن های همپوشانی یک عنصر نسبتاً رایج در سازماندهی ژنوم ویروس ها و احتمالاً پروکاریوت ها هستند. در یوکاریوتها، ناپیوستگی ژن امکان سنتز انواع پپتیدها از یک توالی DNA را نیز فراهم میکند.
با در نظر گرفتن همه این موارد، اصلاح تعریف ژن ضروری است. بدیهی است که دیگر نمیتوانیم درباره ژن به عنوان یک توالی پیوسته از DNA صحبت کنیم که به طور منحصربهفردی یک پروتئین خاص را رمزگذاری میکند. ظاهراً در حال حاضر، فرمول "یک ژن - یک پلی پپتید" هنوز باید قابل قبول ترین باشد، اگرچه برخی از نویسندگان پیشنهاد می کنند که آن را تغییر دهید: "یک پلی پپتید - یک ژن". در هر صورت، اصطلاح ژن باید به عنوان یک واحد عملکردی از مواد ارثی درک شود که به دلیل ماهیت شیمیایی خود یک پلی نوکلئوتید است و امکان سنتز یک زنجیره پلی پپتیدی، tRNA یا rRNA را تعیین می کند.
یک ژن، یک آنزیم.
در سال 1940، J. Beadle و Edward Tatum از یک رویکرد جدید برای مطالعه اینکه چگونه ژن ها متابولیسم را در یک موضوع تحقیقاتی راحت تر - قارچ میکروسکوپی Neurospora crassa - استفاده کردند. هیچ فعالیت آنزیم متابولیک دیگری وجود نداشت. و این منجر به این واقعیت شد که قارچ جهش یافته قادر به سنتز متابولیت خاصی به تنهایی (مثلاً اسید آمینه لوسین) نبود و تنها زمانی می توانست زنده بماند که لوسین به محیط غذایی اضافه شود. نظریه "یک ژن، یک آنزیم" که توسط جی بیدل و ای. تاتوم فرموله شد، به سرعت در میان ژنتیک دانان به رسمیت شناخته شد و خود آنها جایزه نوبل را دریافت کردند.
مواد و روش ها. انتخاب به اصطلاح "جهش های بیوشیمیایی" که منجر به اختلال در عملکرد آنزیم هایی می شود که مسیرهای متابولیک متفاوتی را فراهم می کنند نه تنها برای علم، بلکه برای عمل نیز بسیار مثمر ثمر است. ابتدا به پیدایش ژنتیک و انتخاب میکروارگانیسمهای صنعتی منجر شدند و سپس به صنعت میکروبیولوژیکی که از سویههایی از میکروارگانیسمها استفاده میکنند که مواد استراتژیک مهمی مانند آنتیبیوتیکها، ویتامینها، اسیدهای آمینه و غیره را بیش از حد تولید میکنند. اصول انتخاب و مهندسی ژنتیک. سویه های ابر تولید کننده بر این ایده استوار است که "یک ژن برای یک آنزیم کد می کند." و اگرچه این ایده برای عمل عالی است، سود چند میلیون دلاری به همراه دارد و جان میلیون ها نفر را نجات می دهد (آنتی بیوتیک ها) - نهایی نیست. یک ژن فقط یک آنزیم نیست.
| " |
ژنتیک- علم به هیچ وجه جوان نیست، تحقیقات در مورد آن برای چندین قرن انجام شده است، از مندل در سال 1865 تا به امروز. اصطلاح "ژن" برای یک واحد ویژگی ارثی برای اولین بار توسط یوهانسن در سال 1911 ارائه شد و در دهه 1940 توسط مفهوم "یک ژن، یک آنزیم" که توسط تاتوم و بیدل پیشنهاد شد، اصلاح شد.
این موقعیت در آزمایشات بر روی مگس مگس سرکه مشخص شد، اما به همان اندازه در مورد انسان نیز صدق می کند. در نهایت، زندگی همه موجودات توسط DNA آنها تعیین می شود. مولکول DNA انسان از همه موجودات دیگر بزرگتر است و ساختار آن پیچیده تر است، اما ماهیت عملکرد آن در همه موجودات زنده یکسان است.
مفهوم " یک ژن - یک آنزیم"، که بر اساس ایده های Tatum و Beadle بوجود آمد، می تواند به صورت زیر فرموله شود:
1. همه چیز فرآیندهای بیولوژیکیتحت کنترل ژنتیکی هستند.
2. تمام فرآیندهای بیوشیمیایی به صورت واکنش های مرحله به مرحله رخ می دهد.
3. هر واکنش بیوشیمیایی در نهایت تحت کنترل ژن های مختلف فردی است.
4. جهش در یک ژن خاص منجر به تغییر در توانایی سلول برای انجام یک واکنش شیمیایی خاص می شود.
از آن زمان، مفهوم "یک ژن - یک آنزیم" تا حدودی گسترش یافته است و اکنون به نظر می رسد " یک ژن - یک پروتئین" علاوه بر این، تحقیقات اخیر نشان میدهد که برخی از ژنها برای تولید پروتئینهای منحصربهفرد در هماهنگی با ژنهای دیگر عمل میکنند، بهعنوان مثال، برخی از ژنها ممکن است بیش از یک پروتئین را رمزگذاری کنند.
ژنوم انسانحاوی حدود 3 میلیارد جفت نوکلئوتید است. اعتقاد بر این است که شامل 50000 تا 100000 است. پس از رمزگشایی ژنوم، مشخص شد که تنها حدود 30000 ژن وجود دارد، تعامل این ژن ها بسیار پیچیده تر از حد انتظار است. ژنها در رشتههای DNA کدگذاری میشوند که با پروتئینهای هستهای خاصی ترکیب میشوند و کروموزومها را تشکیل میدهند.
ژن ها- نه فقط بخش های DNA: آنها توسط توالی های کد کننده - اگزون ها، پراکنده با توالی های غیر کد کننده - نیترون ها تشکیل می شوند. اگزون ها، به عنوان بخش بیان شده DNA، تنها بخش کوچکی از مهم ترین مولکول ارگانیسم را تشکیل می دهند. بیشتر آن بیان نمی شود، توسط نیترون ها تشکیل می شود و اغلب به آن DNA "خاموش" می گویند.
اندازه و ساختار تقریبی ژنوم انساندر شکل زیر ارائه شده اند. طول عملکردی کروموزوم انسان بر حسب سانتی مورگانید بیان می شود. سانتی مورگانید (cM) فاصله ای است که احتمال عبور از آن در طول میوز 1٪ است. تجزیه و تحلیل پیوند ژن نشان داده است که طول ژنوم انسان حدود 3000 سانتی متر است.
میانگین کروموزومحاوی تقریباً 1500 ژن است که در 130 میلیون جفت باز نوکلئوتیدی کدگذاری شده اند. شکل زیر به صورت شماتیک اندازه های فیزیکی و عملکردی ژنوم را نشان می دهد: اولی به صورت جفت نوکلئوتیدی و دومی در سانتی متر محاسبه می شود. بیشتر ژنوم انسان با DNA "ساکت" نشان داده می شود و بیان نمی شود.
بر ماتریس DNAدر نتیجه فرآیند رونویسی، RNA و سپس پروتئین سنتز می شود. در نتیجه، توالی DNA به طور کامل توالی پروتئین های عملکردی سلول را تعیین می کند. تمام پروتئین ها به شرح زیر سنتز می شوند:
DNA => RNA => پروتئین

دستگاه ژنتیکی انسان و سایر پستانداران پیچیده تر از سایر موجودات زنده است، زیرا بخش هایی از برخی از ژن ها در پستانداران را می توان با بخش هایی از ژن های دیگر ترکیب کرد. ژن ها، منجر به سنتز یک پروتئین کاملاً جدید یا کنترل یک عملکرد سلولی جداگانه می شود.
در نتیجه، در انسان می توان تعداد ژن های بیان شده را بدون افزایش واقعی حجم ژن های بیان شده افزایش داد. DNAیا تعداد مطلق ژن ها.
به طور کلی، حدود 70٪ از کل مواد ژنتیکی بیان نمی شود.
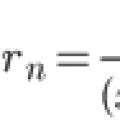 بسط سری تیلور راه حل تقریبی مسئله کوشی برای معمولی
بسط سری تیلور راه حل تقریبی مسئله کوشی برای معمولی چه چیزی در مدرسه آموزش داده نمی شود چه چیزی در مدرسه آموزش داده نمی شود بپرسید
چه چیزی در مدرسه آموزش داده نمی شود چه چیزی در مدرسه آموزش داده نمی شود بپرسید فرمول تفکر پول (A
فرمول تفکر پول (A