Nukleotiidide, nukleiinhapete struktuur ja bioloogiline roll. DNA replikatsioon ja transkriptsioon
Nukleotiidid on nukleosiidide fosforestrid.
Nende keemiline koostis: lämmastikalus (A.O.) + pentoos + fosforhape
Fosforestrid moodustuvad pentooside hüdroksüülrühmade osalusel. Fosforestri rühmade asukohti tähistatakse tavaliselt tähistusega ("), näiteks: 5" , 3 "
Esialgne lühike teave: Nukleotiidid mängivad raku elus äärmiselt olulist rolli.
Nukleotiidide klassifikatsioon
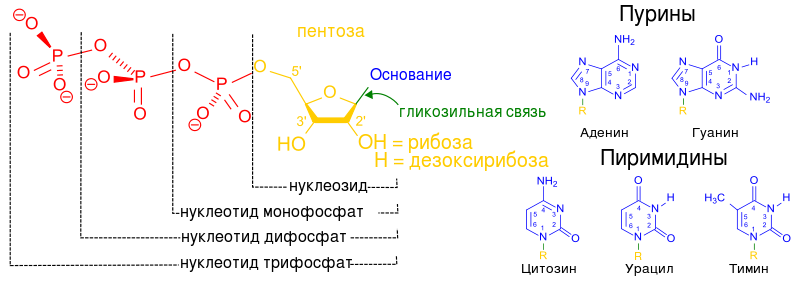
Ühest molekulist koosnevad nukleotiidid A.O, pentoos, fosforhape, kutsutakse mononukleotiidid. Mononukleotiidid võivad sisaldada ühte fosforhappe molekuli, kahte või kolme üksteisega ühendatud fosforhappe molekuli.
Kombinatsioon kaks mononukleotiidi tavaliselt kutsutakse dinukleotiid. IN Dinukleotiid sisaldab tavaliselt erinevaid lämmastiku aluseid või üht muud tsüklilist ühendit, näiteks vitamiini.
Biokeemilistes protsessides mängivad erilist rolli tsüklilised mononukleotiidid.
Mononukleotiidide nomenklatuur.
Pealkirja juurde nukleosiid lisatud fosfaadijääkide koguse alusel, ʼʼ monofosfaatʼʼ, ʼʼdifosfaatʼʼ, "trifosfaat"ʼʼ, mis näitab nende asukohta pentoositsüklis - koha digitaalne tähistamine sümboliga ("),
Fosfaatrühma asukoht positsioonis (5") on kõige levinum ja tüüpilisem, seetõttu võib selle ära jätta (AMP, GTP, UTP, d AMF jne)
Ülejäänud positsioonid tuleb näidata (3" - AMP, 2" - AMP, 3" - d AMF)
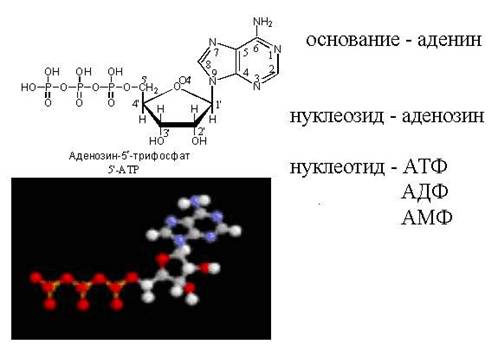
5"-adenosiinmonofosfaat
(5" – AMF või AMF)
Levinumate nukleotiidide nimetused
| nukleosiid | nukleosiidmonofosfaat | nukleosiiddifosfaat | nukleosiidtrifosfaat |
| adenosiin | 5"-adenosiinmonofosfaat (5"-AMP või AMP) 5"-adenüülhape | 5"-adenosiindifosfaat (5"-ADP või ADP) | 5"-adenosiintrifosfaat (5"-ATP või ATP) |
| adenosiin | 3"-adenosiinmonofosfaat (3"-AMP) 3"-adenüülhape | ei leitud in vivo | ei leitud in vivo |
| guanosiin | 5"-guanosiinmonofosfaat (5"-GMP või GMP) | 5"-guanosiindifosfaat (5"-GDP või HDF) | 5"-guanosiintrifosfaat (5"-GTP või GTP) |
| guanosiin | 3"-guanosiinmonofosfaat (3"-GMP) 3"-guanüülhape | ei leitud in vivo | ei leitud in vivo |
| desoksüadenosiin | 5"-desoksüadenosiinmonofosfaat (5"- d AMF või d AMF) | 5"-desoksüadenosiindifosfaat (5"- d ADFili d ADF) | 5"-desoksüadenosiintrifosfaat (5"- d ATFili d ATP) |
| uridiin | 5"-uridiinmonofosfaat (5"-UMP või UMP) | 5"-uridiindifosfaat (5"-UDP või UDF) | 5"-uridiintrifosfaat (5"-UTP või UTP) |
| tsütidiin | 5"-tsütidiinmonofosfaat (5" - CMP või CMP) | 5"-tsütidiindifosfaat (5"-CDP või CDP) | 5"-tsütidiintrifosfaat (5"-CTP või CTP) |
Riboosi osalusel moodustunud nukleotiidid võivad sisaldada fosforhappe jääke kolmes asendis (5", 3", 2") ja desoksüriboosi osalusel - ainult kahes positsioonis (5", 3"), positsioonis 2" seal ei ole hüdroksüülrühm.See asjaolu on DNA struktuuri seisukohalt väga oluline.
Hüdroksürühma puudumisel teises positsioonis on kaks olulist tagajärge:
DNA glükosiidsideme polarisatsioon väheneb ja see muutub hüdrolüüsi suhtes vastupidavamaks.
2-O-desoksüriboos ei saa läbida epimeriseerumist ega muutumist ketoosiks.
Rakus muundatakse nukleosiidmonofosfaat järjestikku difosfaadiks ja seejärel trifosfaadiks.
Näiteks: AMP ---> ADP ---> ATP
Nukleotiidide bioloogiline roll
Kõik nukleosiiddifosfaadid Ja nukleosiidtrifosfaadid kuuluvad suure energiasisaldusega (makroergiliste) ühendite hulka.
Nukleosiidtrifosfaadid osaleda sünteesis nukleiinhapped, tagavad bioorgaaniliste ühendite ja biokeemiliste protsesside aktiveerumise, mis tekivad energia kulutamisel. Adenosiintrifosfaat (ATP) on inimkehas kõige levinum kõrge energiasisaldusega ühend. ATP sisaldus imetajate skeletilihastes on kuni 4 g/kg, üldine sisu umbes 125 ᴦ. Inimestel ulatub ATP metabolismi kiirus 50 kg-ni päevas. ATP hüdrolüüs tekitab adenosiindifosfaat(ADF)
Makroergilised ühendused
ATP sisaldab erinevad tüübid keemilised sidemed:
N-β-glükosiidne
Ester
Kaks anhüdriidi (bioloogiliselt kõrge energiaga)
Tingimustes in vivo kõrge energiaga ATP sideme hüdrolüüsiga kaasneb energia vabanemine (umbes 35 kJ/mol), mis tagab muid energiast sõltuvaid biokeemilisi protsesse.
ATP + H2O - ensüüm ATP hüdrolaas -> ADP + H3 PO4
Vesilahustes ADP ja ATP ebastabiilne . 0 0 juures püsib SATP vees stabiilne vaid paar tundi ja keetmisel 10 minutit.
Leelise mõjul hüdrolüüsuvad kaks terminaalset fosfaati (anhüdriidsidemed) kergesti, kuid viimane (esterside) on raskendatud. Happelise hüdrolüüsi käigus hävib N-glükosiidside kergesti.
Esimest korda vabanes ATP lihastest 1929 ᴦ. K. Loman. Keemiline süntees viidi läbi aastal 1948 ᴦ. A. Todd.
Tsüklilised nukleotiidid on vahendajad hormoonsignaalide edastamisel, muutes ensüümide aktiivsust rakus.
Οʜᴎ moodustuvad nukleosiidtrifosfaatidest.
ATP - tsüklaasensüüm -> cAMP + H4 P2 O7
Pärast toimingu lõppemist toimub tsüklilise nukleotiidi hüdrolüüs. . Võib moodustuda kaks ühendit, 5"-AMP ja 3"-AMP, kuid bioloogilistes tingimustes moodustub ainult 5"-AMP,
Tsükliline adenosiinmonofosfaat (cAMP)
11.5.Nukleiinhapete struktuur
RNA ja DNA esmane struktuur on nukleotiidide järjestikune seos polünukleotiidahelas. Polünukleotiidahela karkass koosneb süsivesikute ja fosfaadi jääkidest, heterotsüklilised lämmastikualused on N-β glükosiidsidemete kaudu ühendatud süsivesikutega. Bioloogilisest vaatenurgast on kõige olulisemad kolmikud – nukleotiidide plokid, mis koosnevad kolmest lämmastikualusest, millest igaüks kodeerib aminohapet või omab spetsiifilist signaalimisfunktsiooni.
NC struktuuri saab skemaatiliselt kujutada:
5" 3" 5" 3" 5" 3"
fosfaat -- pentoos -- fosfaat -- pentoos -- fosfaat -- pentoos -OH
DNA esmases struktuuris Alusta ahelad määratakse pentoosiga, mis sisaldab 5. positsioonis fosfaati. Polünukleotiidahela pentoosid on ühendatud fosfaatsidemetega 3 "→ 5". Peal lõpp ahelad positsioonis 3"-pentoos OH-rühm jääb vabaks.
Kõrgemat järku DNA struktuur – topeltheeliks
Teaduslik kirjeldus sekundaarne struktuur DNA viitab suurimad avastused inimkond kahekümnendal sajandil. Biokeemik D. Watson ja füüsik F. Crick 1953. aastal pakkusid nad välja DNA struktuuri mudeli ja replikatsiooniprotsessi mehhanismi. 1962. aastal. pälvisid nad Nobeli preemia.
Populaarsel kujul on lugu kirjeldatud James Watsoni raamatus "The Double Helix", M.: Mir, 1973. Raamat kirjeldab väga huvitavalt koostöö ajalugu, huumori ja autori kerge irooniaga sellise suhtes märkimisväärne sündmus, mille õnnelikud “süüdlased” olid kaks noort teadlast. Alates DNA struktuuri avastamisest on inimkond saanud tööriista uue suuna arendamiseks - biotehnoloogia, valkude süntees geenide rekombinatsiooni teel (meditsiinitööstuses toodavad hormoone insuliin, erütropoetiin ja paljud teised).
Teadusuuringud aitasid kaasa DNA struktuuri avastamisele E. Chargaffa DNA keemilise koostise kohta. Ta sai teada:
Pürimidiini aluste arv on võrdne puriini aluste arvuga
Tümiini kogus on võrdne adeniini kogusega ja tsütosiini kogus on võrdne
A = T G = C
A + G = T + C
A + C = T + G
Seda suhet nimetatakse Chargaffi reeglid .
DNA molekul koosneb kahest keerdunud heeliksist. Iga heeliksi skelett on vahelduvate desoksüriboosi ja fosforhappe jääkide ahel. Spiraalid on orienteeritud nii, et need moodustavad kaks ebavõrdset spiraalset soont, mis kulgevad paralleelselt peateljega. Need sooned on täidetud valkudega histoonid. Lämmastikalused asuvad spiraali sees, peaaegu risti peateljega ja moodustavad ahelate vahel komplementaarsed paarid A...T ja G...C.
Iga raku DNA molekulide kogupikkus ulatub 3 cm-ni, raku läbimõõt on keskmiselt 10–5 m, DNA läbimõõt vaid 2 ‣‣‣10–9 m.
Topeltheeliksi põhiparameetrid:
* läbimõõt 1,8-2nm,
* üks pööre sisaldab 10 nukleotiidi
* pöörde sammu kõrgus ~ 3,4 nm
* kahe nukleotiidi vaheline kaugus on 0,34 nm.
Alused asetsevad keti teljega risti.
* polünukleotiidahelate suunad on antiparalleelsed
* seos desoksüriboosi furanoositsüklite vahel
fosforhape viiakse läbi positsioonist 3 "asendisse 5" sisse
iga vooluringi.
* Ahela algus – pentoosi hüdroksüülrühm fosforüülitakse positsioonis
5`, ahela ots on pentoosi vaba hüdroksüülrühm asendis 3`.
* DNA-s ja RNA-s on nukleosiidifragmendid antikonformatsioonis, puriini pürimidiinitsükkel asub glükosiidsidemest paremal. Ainult see asend võimaldab moodustada komplementaarse paari (vt nukleotiidide valemeid)
* Lämmastikualuste vahel esineb kolme tüüpi interaktsioone:
1. “Riik”, kaasatud on kahe ahela üksteist täiendavad paarid. Kahe lämmastikaluse (T – A, U – C) vahel toimub elektronide “tsükliline” ülekanne, tekib täiendav p-elektronsüsteem, mis tagab täiendava interaktsiooni ja kaitseb lämmastikaluseid soovimatute keemiliste mõjude eest. vahel adeniini ja tümiini vahel luuakse kaks vesiniksidet ning guaniini ja tsütosiini vahel kolm vesiniksidet.
2. “Vertikaalne” (virnastamine), virnastamise tõttu on kaasatud ühe ahela lämmastikualused. "Vinastamise interaktsioon" isegi on rohkem tähtsus struktuuri stabiliseerimisel kui vastastikmõju täiendavates paarides
3. Koostoime veega mängib olulist rolli kaksikheeliksi ruumilise struktuuri säilitamisel, mis võtab kõige kompaktsema struktuuri, et vähendada veega kokkupuutepinda ja suunab hüdrofoobsed heterotsüklilised alused spiraali.
Nukleoproteiinikomplekside struktuur ja koostis
Nukleiinhappe seondumisel valguga osalevad mitut tüüpi interaktsioonid:
Elektrostaatiline
Vesiniksidemed
Hüdrofoobne
Röntgenstruktuurianalüüsi tulemuste põhjal konstrueeriti arvutimodelleerimise abil DNA, ribosoomide, infoosoomide ja viiruslike nukleiinhapete reaalsed kolmemõõtmelised mudelid.
Histooni DNA valkudel on selged põhiomadused ja need eristuvad kõrge evolutsioonilise säilivuse poolest. Kahe aluselise aminohappe lüsiini/arginiini suhte alusel jagatakse need 5 klassi: H1, H2A, H 2B, H3, H4
Nukleotiidid – mõiste ja liigid. Kategooria "Nukleotiidid" klassifikatsioon ja tunnused 2017, 2018.
- need on komplekssed monomeerid, millest on kokku pandud heteropolümeeri molekulid. DNA ja RNA. Vabad nukleotiidid osalevad elu signaali- ja energiaprotsessides. DNA nukleotiididel ja RNA nukleotiididel on ühine struktuuriplaan, kuid need erinevad pentoossuhkru struktuuri poolest. DNA nukleotiidid kasutavad suhkru desoksüriboosi, RNA nukleotiidid aga riboosi.
Nukleotiidi struktuur
Iga nukleotiidi võib jagada kolmeks osaks:
1. Süsivesik on viieliikmeline pentoossuhkur (riboos või desoksüriboos).
2. Fosforijääk (fosfaat) on fosforhappe jääk.
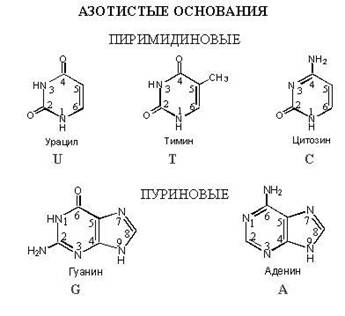
3. Lämmastikalus on ühend, mis sisaldab palju lämmastikuaatomeid. Nukleiinhapped kasutavad ainult 5 tüüpi lämmastiku aluseid: adeniin, tümiin, guaniin, tsütosiin, uratsiil. DNA-s on 4 tüüpi: adeniin, tümiin, guaniin, tsütosiin. RNA-s on ka 4 tüüpi: Adeniin, Uratsiil, Guaniin, Tsütosiin.On lihtne märgata, et RNA-s on tümiin asendatud Uratsiiliga võrreldes DNA-ga.
Kindral struktuurvalem pentoos (riboos või desoksüriboos), mille molekulid moodustavad nukleiinhapete "skeleti":
Kui X asendatakse H-ga (X = H), saadakse desoksüribonukleosiidid; kui X on asendatud OH-ga (X = OH), siis saadakse ribonukleosiidid. Kui asendate R asemel lämmastikaluse (puriin või pürimidiin), saate konkreetse nukleotiidi.
Oluline on pöörata tähelepanu süsinikuaatomite positsioonidele pentoosis, mis on tähistatud kui 3" ja 5". Süsinikuaatomite nummerdamine algab ülaosas olevast hapnikuaatomist ja läheb päripäeva. Viimane on süsinikuaatom (5"), mis asub väljaspool pentoositsüklit ja moodustab, võib öelda, pentoosi "saba". Nii et nukleotiidide ahela kasvatamisel suudab ensüüm kinnitada ainult uue nukleotiidi süsinikule 3" ja mitte ühelegi teisele . Seetõttu ei saa nukleotiidahela 5-tollist otsa kunagi jätkata; pikendada saab ainult 3-tollist otsa.


Võrrelge RNA nukleotiidi DNA nukleotiidiga.
Proovige välja selgitada, mis nukleotiid see on selles esituses:

ATP - vaba nukleotiid
cAMP on "silmus" ATP molekul

Nukleotiidi struktuuri diagramm
Pange tähele, et aktiveeritud nukleotiidil, mis on võimeline pikendama DNA või RNA ahelat, on "trifosfaadi saba". Just selle "energiaga küllastunud" sabaga saab see liituda olemasoleva kasvava nukleiinhappe ahelaga. Fosfaadi saba asub 5. süsinikul, nii et see süsiniku asend on juba fosfaatide poolt hõivatud ja on seal kinnitumiseks. Mille külge peaksin selle kinnitama? Ainult süsinikule positsioonis 3". Kui antud nukleotiid on kinnitunud, muutub see ise sihtmärgiks järgmise nukleotiidi kinnitumisel. "Vastuvõttev pool" annab süsiniku positsioonis 3" ja "saabuv pool" klammerdub selle külge fosfaadi saba asub asendis 5". Üldiselt kasvab kett 3" küljelt.
DNA nukleotiidahela pikendamine
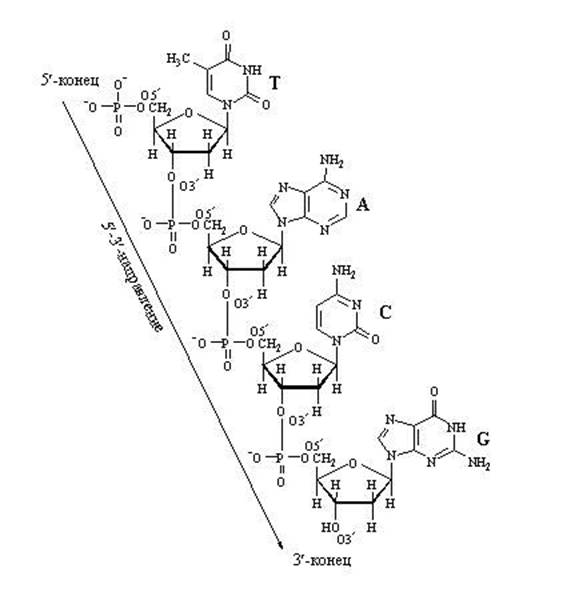
Nukleotiidide vahelistest "pikisuunalistest" sidemetest tulenev ahela pikendus võib minna ainult ühes suunas: 5" ⇒ kuni 3", sest uut nukleotiidi saab lisada ainult ahela 3" otsa, kuid mitte 5" otsa.
Nukleotiidide paarid, mis on ühendatud nende lämmastikualuste komplementaarsete ristsidemetega
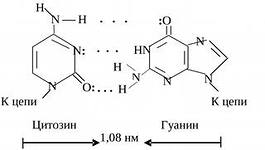

DNA topeltheeliksi piirkond

Leidke märke, et kaks DNA ahelat on paralleelsed.
Leidke topelt- ja kolmiksidemetega nukleotiidipaarid.
Nukleosiidid on omakorda N-glükosiidid, mis sisaldavad heterotsüklilist osa, mis on lämmastikuaatomi kaudu seotud suhkrujäägi C-1 aatomiga.
Looduses on levinumad nukleotiidid puriinide või pürimidiinide β -N-glükosiidid ja pentoosid - D-riboos või D-2-desoksüriboos. Sõltuvalt pentoosi struktuurist eristatakse ribonukleotiide ja desoksüribonukleotiide, mis on komplekssete bioloogiliste polümeeride molekulide monomeerid. (polünukleotiidid)- vastavalt RNA või DNA.
Nukleotiidides sisalduv fosfaadijääk moodustab tavaliselt estersideme ribonukleosiidide 2'-, 3'- või 5'-hüdroksüülrühmadega, 2'-desoksünukleosiidide puhul on 3'- või 5'-hüdroksüülrühmad esterdatud.
Enamik nukleotiide on ortofosforhappe monoestrid, kuid teada on ka nukleotiidide diestreid, milles on esterdatud kaks hüdroksüüljääki – näiteks tsüklilised nukleotiidid tsükloadeniin ja tsükloguaniinmonofosfaadid (cAMP ja cGMP). Koos nukleotiididega on looduses levinud ka ortofosforhappe estrid (monofosfaadid), pürofosforhappe mono- ja diestrid (difosfaadid, näiteks adenosiindifosfaat) ja tripolüfosforhappe monoestrid (trifosfaadid, näiteks adenosiintrifosfaat).
Nomenklatuur
| Kood | Tähendab | Täiendav paar |
|---|---|---|
| A | T DNA-s; U RNA-s |
|
| C | G | |
| G | C | |
| või |
T DNA-s; U RNA-s |
A |
| M | A või C |
K |
| R | A või G |
Y |
| W | A või T |
W |
| S | C või G |
S |
| Y | C või T |
R |
| K | G või T |
M |
| V | A või C või G |
B |
| H | A või C või T |
D |
| D | A või G või T |
H |
| B | C või G või T |
V |
| X või N |
A või C või G või T(U) |
ükskõik milline |
Kahest nukleotiidmolekulist koosnevaid ühendeid nimetatakse dinukleotiidid, kolmest - trinukleotiidid, väikesest arvust - oligonukleotiidid ja paljudelt - polünukleotiidid või nukleiinhapped.
Nukleotiidide nimetused on lühendid standardsete kolme- või neljatäheliste koodide kujul.
Kui lühend algab väikese tähega "d" (ing. d), mis tähendab desoksüribonukleotiidi; tähe "d" puudumine tähendab ribonukleotiidi. Kui lühend algab väikese tähega "ts" (ing. c), mis tähendab, et me räägime nukleotiidi tsüklilisest vormist (näiteks cAMP).
Lühendi esimene suurtäht tähistab konkreetset lämmastikualust või võimalike nukleiinsete aluste rühma, teine täht näitab fosforhappejääkide arvu struktuuris (M - mono-, D - di-, T - tri-) ja kolmas suurtäht on alati täht F ("-phosphate"; inglise keel. P).
Nukleiinaluste ladina ja vene koodid:
- T - T: tümiin (5-metüüluratsiil), mida leidub DNA bakteriofaagides, asendab RNA-s uratsiili;
- U - U: RNA-s leiduv uratsiil asendab DNA-s tümiini.
Nukleotiidsete aluste tähistamiseks kasutatavad üldtunnustatud tähekoodid vastavad Rahvusvahelise Puhta ja Rakenduskeemia Liidu (lühendatult inglise keeles) vastu võetud nomenklatuurile. IUPAC, IUPAC) ja Rahvusvaheline Biokeemia ja Molekulaarbioloogia Liit (Inglise)vene keel, lühendatult - inglise keel. IUBMB). Kui DNA või RNA järjestuse sekveneerimisel tekib kahtlus konkreetse nukleotiidi määramise täpsuses, kasutatakse lisaks viiele peamisele nukleotiidile (A, C, T, G, U) ka teisi ladina tähestiku tähti. millised võivad olla kõige tõenäolisemad nukleotiidid antud järjestuse positsioonides. Samu lisatähti kasutatakse degenereerunud positsioonide tähistamiseks (mis ei lange erinevates homoloogsetes järjestustes kokku), näiteks PCR-i praimerite järjestuse kirjutamisel.
Sekveneeritud DNA lõikude (geen, sait, kromosoom) või kogu genoomi pikkus on näidatud nukleotiidipaaridena (bp) või aluspaaridena (eng. aluspaarid, lühendatult bp), mis tähendab selle all kaheahelalise nukleiinhappemolekuli elementaarüksust, mis koosneb kahest paaritud komplementaarsest alusest.
Bioloogiline roll
1. Universaalne energiaallikas (ATP ja selle analoogid).
2. Need on monomeeride aktivaatorid ja kandjad rakus (UDP-glükoos)
6. Need on nukleiinhapete koostises olevad monomeerid, mis on ühendatud 3'-5'-fosfodiestersidemetega.
Lugu
Premolekulaarses geneetikas kasutati spetsiaalset terminit DNA struktuuri väikseima elemendi tähistamiseks, mis võib olla spontaanse või indutseeritud mutatsiooni all. recon. Nüüd on näidatud, et väikseim element on üks nukleotiid (või üks lämmastikualus nukleotiidis), mistõttu seda terminit enam ei kasutata. Mõiste defineerimiseks mutatsiooniüksus terminit kasutati Mouton. Nüüd on näidatud, et mutatsioon võib avalduda fenotüüpiliselt isegi siis, kui asendatakse üks nukleotiid (või lämmastikalus nukleotiidis), seega vastab termin muton ühele nukleotiidile.
Kirjutage ülevaade artiklist "Nukleotiidid"
Märkmed
Vaata ka
Lingid
- Nukleotiidid- artikkel Suurest Nõukogude Entsüklopeediast.
- Nukleotiidsete aluste tähtkoodid vastavalt IUPAC ja IUBMB nomenklatuurile (Inglise)vene keel
- veebiajakirjas
|
||||||||||||||||||||||||||||||||||
Nukleotiide iseloomustav väljavõte
Kasakat kutsuti ja küsitleti; Kasakate komandörid tahtsid kasutada võimalust hobuste tagasipüüdmiseks, kuid üks armee kõrgeimaid auastmeid tundev komandör teatas sellest staabikindralile. Viimasel ajal on olukord armee peakorteris olnud äärmiselt pingeline. Ermolov palus paar päeva varem Bennigseni jõudes, et ta kasutaks oma mõju ülemjuhatajale, et alustada pealetungi."Kui ma ei teaks sind, siis ma arvaks, et sa ei taha seda, mida sa palud." "Niipea, kui ma üht asja annan, teeb Tema rahulik Kõrgus tõenäoliselt vastupidist," vastas Bennigsen.
Kasakate uudised, mida saadetud patrullid kinnitasid, tõestasid sündmuse lõplikku küpsust. Venitatud keel hüppas, kell susises ja kellamäng hakkas mängima. Vaatamata kogu oma väljamõeldud jõule, intelligentsusele, kogemustele, inimeste tundmisele, võttis Kutuzov, võttes arvesse suveräänile isiklikult aruandeid saatnud Bennigseni märkust, sama soovi, mida väljendasid kõik kindralid, tema enda võetud suverääni soovi. ja kasakate kokkutoomine, ei suutnud enam pidurdada vältimatut liikumist ja andis korraldusi selleks, mida ta pidas kasutuks ja kahjulikuks – ta õnnistas saavutatud fakti.
Bennigseni esitatud noot pealetungi vajalikkuse kohta ja kasakate teave prantslaste katmata vasaku tiiva kohta olid vaid viimased märgid vajadusest pealetungi tellida ning pealetung oli kavandatud 5. oktoobriks.
4. oktoobri hommikul kirjutas Kutuzov käsukorraldusele alla. Tol luges selle Jermolovile ette, kutsudes teda edasisi korraldusi täitma.
"Olgu, okei, mul pole praegu aega," ütles Ermolov ja lahkus onnist. Toli koostatud dispositsioon oli väga hea. Nii nagu Austerlitzi dispositsioonis, kirjutati, kuigi mitte saksa keeles:
"Die erste Colonne marschiert [Esimene veerg läheb (saksa)] seda ja teist, die zweite Colonne marschiert [teine veerg läheb (saksa keel)] siia ja sinnapoole" jne. Ja kõik need veerud paberil jõudsid määratud ajal oma kohale ja hävitas vaenlase. Kõik oli, nagu kõigis dispositsioonides, suurepäraselt läbi mõeldud ja nagu kõigis dispositsioonides, ei saabunud ükski kolonn omal ajal ja oma kohale.
Kui dispositsioon oli vajalikus arvus eksemplaris valmis, kutsuti kohale ohvitser, kes saadeti Ermolovi juurde, et anda talle paberid täitmiseks. Noor ratsaväeohvitser, Kutuzovi korrapidaja, olles rahul talle antud ülesande tähtsusega, läks Ermolovi korterisse.
"Oleme lahkunud," vastas Jermolov korrapidaja. Ratsaväeohvitser läks kindrali juurde, kes külastas sageli Ermolovit.
- Ei ja kindralit pole.
Ratsaväeohvitser, istus hobuse seljas, ratsutas teise juurde.
- Ei, nad lahkusid.
“Kuidas ma ei vastuta hilinemise eest! Milline häbi! - mõtles ohvitser. Ta tegi ringkäigu kogu laagris. Mõni ütles, et nägi Ermolovit koos teiste kindralitega kuhugi minemas, mõni ütles, et ilmselt on ta jälle kodus. Ametnik otsis lõunat söömata kuni kella kuueni õhtul. Jermolovit polnud kuskil ja keegi ei teadnud, kus ta on. Ohvitser sõi ruttu seltsimehega näksi ja läks tagasi avangardi Miloradovitši juurde. Ka Miloradovitšit polnud kodus, kuid siis öeldi talle, et Miloradovitš oli kindral Kikini ballil ja ka Jermolov peab seal olema.
- Kus see on?
"Seal, Echkinos," ütles kasakate ohvitser, osutades kaugel asuvale maaomanikule.
- Kuidas seal keti taga on?
- Nad saatsid kaks meie rügementi ketti, seal toimub praegu selline lõbustus, see on katastroof! Kaks muusikat, kolm laulukirjutajate koori.
Ohvitser läks keti taha Echkini juurde. Kaugelt majale lähenedes kuulis ta sõduri tantsulaulu sõbralikke, rõõmsaid helisid.
"Niitudel, ah... heinamaadel!.." - kuulis ta teda vilistamas ja kõlksumas, aeg-ajalt häälte kisa alla uppumas. Ohvitser tundis nendest helidest hinges rõõmu, kuid samas kartis, et on ise süüdi, et talle usaldatud tähtsat käsku nii kaua edasi ei andnud. Kell oli juba üheksa. Ta astus hobuse seljast maha ja sisenes venelaste ja prantslaste vahel asuva suure terve mõisahoone veranda ja esikusse. Sahvris ja koridoris askeldasid jalamehed veinide ja roogadega. Akende all olid lauluraamatud. Ohvitser juhatati uksest sisse ja ta nägi järsku koos kõiki armee tähtsamaid kindraleid, sealhulgas suurt, märgatavat Ermolovi kuju. Kõik kindralid olid lahti nööbitavates kitlites, punaste, elavate nägudega ja naersid valju häälega, seisid poolringis. Keset saali meisterdas nutikalt ja osavalt thrasherit üks nägus punakas näoga lühike kindral.
- Ha, ha, ha! Oh jah, Nikolai Ivanovitš! ha, ha, ha!..
Ohvitser tundis, et sel hetkel tähtsa käsuga sisenedes on ta kahekordselt süüdi ja ta tahab oodata; kuid üks kindralitest nägi teda ja, olles teada saanud, milleks ta oli, rääkis Ermolovile. Kortsutatud näoga Ermolov läks ohvitseri juurde ja võttis pärast ärakuulamist temalt paberi, midagi ütlemata.
- Kas sa arvad, et ta lahkus kogemata? - ütles staabikaaslane tol õhtul ratsaväeohvitserile Ermolovi kohta. - Need on asjad, see kõik on meelega. Andke Konovnitsõnile sõit. Vaata, milline jama homme on!
Järgmisel päeval varahommikul tõusis vaoshoitud Kutuzov püsti, palvetas jumalat, riietus ning ebameeldivas teadvuses, et peab juhtima lahingut, mida ta heaks ei kiidanud, istus vankrisse ja sõitis Letashevkast välja. , viis miili Tarutini taga, kohta, kus tuli kokku panna edasiliikuvad kolonnid. Kutuzov ratsutas, jäi magama ja ärkas ja kuulas, kas paremalt on kaadreid, kas asjad hakkavad? Kuid kõik oli endiselt vaikne. Niiske ja pilvise sügispäeva koit alles algas. Tarutinile lähenedes märkas Kutuzov ratsaväelasi, kes viisid oma hobused vette üle tee, mida mööda vanker liikus. Kutuzov vaatas neid lähemalt, peatas vankri ja küsis, mis rügement? Ratsaväelased olid kolonnist, mis oleks pidanud varitsuses olema kaugel ees. "See võib olla viga," arvas vana ülemjuhataja. Kuid veelgi kaugemale sõitnud, nägi Kutuzov jalaväerügemente, relvi kastides, sõdureid pudru ja küttepuudega, aluspükstes. Kutsuti ohvitser. Ametnik teatas, et liikumiskäsku polnud.
"Kuidas sa ei saaks..." alustas Kutuzov, kuid vaikis kohe ja käskis vanemohvitseri enda juurde kutsuda. Olles vankrist välja tulnud, pea maas ja raskelt hingates, vaikselt oodates, kõndis ta edasi-tagasi. Kui kohale ilmus nõutud kindralstaabi ohvitser Eichen, muutus Kutuzov lillaks, mitte sellepärast, et see ohvitser oleks süüdi eksinud, vaid sellepärast, et ta oli väärt teema viha väljendamiseks. Ja värisedes, hingeldades tungis vanamees sellesse raevuseisundisse, millesse ta suutis sattuda, kui ta vihast maas veeres, ründas Eichenit, ähvardades kätega, karjudes ja vulgaarselt vandudes. sõnad. Sama saatus tabas teist kohale tulnud inimest, kapten Brozinit, kes oli milleski süütu.
Kogu elu planeedil koosneb paljudest rakkudest, mis säilitavad tuumas sisalduva geneetilise teabe tõttu oma organisatsiooni korrastatuse. Seda säilitavad, rakendavad ja edastavad komplekssed kõrgmolekulaarsed ühendid - nukleiinhapped, mis koosnevad monomeersetest ühikutest - nukleotiididest. Nukleiinhapete rolli ei saa ülehinnata. Nende struktuuri stabiilsus määrab organismi normaalse funktsioneerimise ning kõik kõrvalekalded struktuuris toovad paratamatult kaasa muutusi rakukorralduses, füsioloogiliste protsesside aktiivsuses ja rakkude elujõulisuses üldiselt.
Nukleotiidi mõiste ja selle omadused
Iga RNA on kokku pandud väiksematest monomeersetest ühenditest – nukleotiididest. Teisisõnu on nukleotiid ehitusmaterjal nukleiinhapetele, koensüümidele ja paljudele teistele bioloogilistele ühenditele, mis on rakule elu jooksul hädavajalikud.
Nende oluliste ainete peamised omadused on järgmised:
Teabe salvestamine ja päritud tunnuste kohta;
. kontrolli teostamine kasvu ja paljunemise üle;
. osalemine ainevahetuses ja paljudes teistes rakus toimuvates füsioloogilistes protsessides.
Nukleotiididest rääkides ei saa jätta peatumata nii olulisel teemal nagu nende struktuur ja koostis.

Iga nukleotiid koosneb:
Suhkru jäägid;
. lämmastikalus;
. fosfaatrühm või fosforhappejääk.
Võime öelda, et nukleotiid on keeruline orgaaniline ühend. Sõltuvalt lämmastikaluste liigilisest koostisest ja pentoosi tüübist nukleotiidstruktuuris jagatakse nukleiinhapped:
desoksüribonukleiinhape või DNA;
. ribonukleiinhape ehk RNA.
Nukleiinhappe koostis
Nukleiinhapetes esindab suhkrut pentoos. See on viiesüsinikuline suhkur, mida DNA-s nimetatakse desoksüriboosiks ja RNA-s riboosiks. Igas pentoosi molekulis on viis süsinikuaatomit, neist neli koos hapnikuaatomiga moodustavad viieliikmelise tsükli ja viies on osa HO-CH2 rühmast.
Iga süsinikuaatomi asukoht pentoosi molekulis on tähistatud araabia numbriga koos algarvuga (1C´, 2C´, 3C´, 4C´, 5C´). Kuna kõigil nukleiinhappemolekulist lugemise protsessidel on range suund, on süsinikuaatomite nummerdamine ja nende asukoht ringis omamoodi õige suuna indikaator.
Hüdroksüülrühmas on fosforhappejääk seotud kolmanda ja viienda süsinikuaatomiga (3C´ ja 5C´). See määrab DNA ja RNA keemilise kuuluvuse hapete rühma.
Lämmastikalus on seotud suhkrumolekuli esimese süsinikuaatomiga (1C').
Lämmastikaluste liigiline koostis
Lämmastikalusel põhinevad DNA nukleotiidid on esindatud nelja tüüpi:
adeniin (A);
. guaniin (G);
. tsütosiin (C);
. tümiin (T).
Esimesed kaks kuuluvad puriinide klassi, kaks viimast kuuluvad pürimidiinide klassi. Molekulmassi poolest on puriinid alati raskemad kui pürimidiinid.

Lämmastikalusel põhinevad RNA nukleotiidid on esindatud:
adeniin (A);
. guaniin (G);
. tsütosiin (C);
. uratsiil (U).
Uratsiil, nagu tümiin, on pürimidiini alus.
Teaduskirjanduses võib sageli leida lämmastikualuste jaoks veel ühe nimetuse - ladina tähtedega (A, T, C, G, U).
Vaatleme üksikasjalikumalt puriinide ja pürimidiinide keemilist struktuuri.

Pürimidiinid, nimelt tsütosiin, tümiin ja uratsiil, koosnevad kahest lämmastikuaatomist ja neljast süsinikuaatomist, moodustades kuueliikmelise ringi. Igal aatomil on oma arv vahemikus 1 kuni 6.
Puriinid (adeniin ja guaniin) koosnevad pürimidiinist ja imidasoolist või kahest heterotsüklist. Puriini alusmolekuli esindavad neli lämmastikuaatomit ja viis süsinikuaatomit. Iga aatom on nummerdatud 1 kuni 9.
Lämmastikaluse ja pentoosijäägi koosmõjul tekib nukleosiid. Nukleotiid on nukleosiidi ja fosfaatrühma ühend.
Fosfodiestersidemete moodustumine
Oluline on mõista küsimust, kuidas nukleotiidid on ühendatud polüpeptiidahelaks ja moodustavad nukleiinhappemolekuli. See juhtub nn fosfodiestersidemete tõttu.
Kahe nukleotiidi vastasmõju tekitab dinukleotiidi. Uue ühendi moodustumine toimub kondenseerumisel, kui ühe monomeeri fosfaadijäägi ja teise monomeeri pentoosi hüdroksürühma vahel tekib fosfodiesterside.
Polünukleotiidide süntees on selle reaktsiooni korduv kordamine (mitu miljonit korda). Polünukleotiidahel ehitatakse üles fosfodiestersidemete moodustumisega suhkrute kolmanda ja viienda süsiniku (3C´ ja 5C´) vahel.
Polünukleotiidide kokkupanek on keeruline protsess, mis toimub ensüümi DNA polümeraasi osalusel, mis tagab ahela kasvu ainult ühest otsast (3´), millel on vaba hüdroksürühm.
DNA molekuli struktuur
DNA molekulil, nagu ka valkudel, võib olla primaarne, sekundaarne ja tertsiaarne struktuur.

Nukleotiidide järjestus DNA ahelas määrab selle primaarse, see moodustub vesiniksidemete tõttu, mille aluseks on komplementaarsuse põhimõte. Teisisõnu, kaksikahela sünteesi ajal kehtib teatud muster: ühe ahela adeniin vastab teise tümiinile, guaniin tsütosiinile ja vastupidi. Adeniini ja tümiini või guaniini ja tsütosiini paarid moodustuvad esimesel kahel ja viimasel juhul kolmel vesiniksidemetel. Selline nukleotiidide ühendus tagab ahelate tugeva ühenduse ja nendevahelise võrdse vahemaa.
Teades ühe DNA ahela nukleotiidjärjestust, saab teise ahela täiendada komplementaarsuse või liitmise põhimõttel.
DNA tertsiaarne struktuur moodustub keeruliste kolmemõõtmeliste sidemete tõttu, mis muudab selle molekuli kompaktsemaks ja mahutab väikesesse rakumahu. Näiteks E. coli DNA pikkus on üle 1 mm, samas kui raku pikkus on alla 5 mikroni.
Nukleotiidide arv DNA-s, nimelt nende kvantitatiivne suhe, järgib Chergaffi reeglit (puriini aluste arv on alati võrdne pürimidiini aluste arvuga). Nukleotiidide vaheline kaugus on konstantne väärtus, mis võrdub 0,34 nm, nagu ka nende molekulmass.
RNA molekuli struktuur
RNA-d esindab üks polünukleotiidahel, mis on moodustatud pentoosi (antud juhul riboosi) ja fosfaadijäägi vahel. See on palju lühem kui DNA. Samuti on erinevusi nukleotiidi lämmastikaluste liigilises koostises. RNA-s kasutatakse pürimidiinaluse tümiini asemel uratsiili. Sõltuvalt kehas täidetavatest funktsioonidest võib RNA-d olla kolme tüüpi.

Ribosomaalne (rRNA) - sisaldab tavaliselt 3000 kuni 5000 nukleotiidi. Nagu vajatud struktuurne komponent osaleb ribosoomide aktiivse tsentri moodustamises, mis on raku ühe olulisema protsessi - valgu biosünteesi koht.
. Transport (tRNA) - koosneb keskmiselt 75–95 nukleotiidist, viib soovitud aminohappe ribosoomi polüpeptiidi sünteesi piirkonda. Igal tRNA tüübil (vähemalt 40) on oma ainulaadne monomeeride või nukleotiidide järjestus.
. Informatsioon (mRNA) – nukleotiidide koostiselt väga mitmekesine. Kannab geneetilise informatsiooni DNA-st ribosoomidesse ja toimib valgumolekulide sünteesi maatriksina.
Nukleotiidide roll organismis
Nukleotiidid rakus täidavad mitmeid olulisi funktsioone:
Kasutatakse nukleiinhapete ehitusplokkidena (puriini ja pürimidiini seeria nukleotiidid);
. osaleda paljudes raku ainevahetusprotsessides;
. on osa ATP-st - rakkude peamisest energiaallikast;
. toimivad redutseerivate ekvivalentide kandjatena rakkudes (NAD+, NADP+, FAD, FMN);
. täidavad bioregulaatorite funktsiooni;
. võib pidada ekstratsellulaarse regulaarse sünteesi teiseks sõnumitoojaks (näiteks cAMP või cGMP).
Nukleotiid on monomeerne üksus, mis moodustab keerukamaid ühendeid - nukleiinhappeid, ilma milleta on geneetilise teabe edastamine, selle säilitamine ja paljundamine võimatu. Vabad nukleotiidid on peamised signaali- ja energiaprotsesside komponendid, mis toetavad rakkude ja kogu organismi normaalset talitlust.
Inimese kehas on suur hulk orgaanilised ühendid, ilma milleta on võimatu ette kujutada ainevahetusprotsesside stabiilset kulgu, mis toetab kõigi elu. Üks neist ainetest on nukleotiidid – need on nukleosiidide fosforestrid, mis mängivad olulist rolli infoandmete edastamisel, samuti keemilised reaktsioonid rakusisese energia vabanemisega.
Iseseisvate orgaaniliste üksustena moodustavad nad kõigi nukleiinhapete ja enamiku koensüümide täitekoostise. Vaatame lähemalt, mis on nukleosiidfosfaadid ja millist rolli nad inimkehas mängivad.
Millest koosneb aine nukleotiid? Seda peetakse äärmiselt estriks, mis kuulub fosforhapete ja nukleosiidide rühma, mis oma biokeemiliste omaduste poolest on klassifitseeritud N-glükosiidideks ja sisaldavad glükoosi molekulide ja lämmastikuaatomiga seotud heterotsüklilisi fragmente.
Looduses on kõige levinumad nukleotiidid DNA.
Lisaks eristavad nad ka orgaaniline aine sarnaste struktuuriomadustega: ribonukleotiidid, samuti desoksüribonukleotiidid. Kõik need on eranditult monomeersed molekulid, mis kuuluvad keeruka struktuuriga polümeeri tüüpi bioloogilistesse ainetesse.
Neist moodustub kõigi elusolendite RNA ja DNA, alates kõige lihtsamatest mikroorganismidest ja viirusinfektsioonidest kuni inimkehani välja.
Ülejäänud osa fosfori molekulaarstruktuurist nukleosiidfosfaatide hulgas moodustab estersideme kahe, kolme ja mõnel juhul viie hüdroksüülrühmaga korraga. Peaaegu kõik nukleotiidid on eranditult olulised ained, mis moodustuvad ortofosforhappe jääkidest, seetõttu on nende sidemed stabiilsed ega lagune sise- ja väliskeskkonna ebasoodsate tegurite mõjul.
Märge! Nukleotiidide struktuur on alati keeruline ja põhineb monoestritel. Nukleotiidjärjestus võib stressifaktorite mõjul muutuda.
Bioloogiline roll
Nukleotiidide mõju kõigi elusolendite kehas toimuvate protsesside kulgemisele uurivad teadlased, kes uurivad molekulaarne struktuur rakusisene ruum.
Üle maailma teadlaste aastatepikkuse töö tulemuste põhjal saadud laboratoorsete leidude põhjal eristatakse nukleosiidfosfaatide järgmist rolli:
- universaalne elutähtsa energia allikas, mille tõttu rakke toidetakse ja vastavalt hoitakse normaalne töö kuded, mis moodustavad siseorganeid, bioloogilisi vedelikke, epiteeli katet ja veresoonte süsteemi;
- on glükoosmonomeeride transportijad mis tahes tüüpi rakkudes (see on üks süsivesikute metabolismi vorme, mille tarbimisel muundub suhkur seedeensüümide mõjul glükoosiks, mis kantakse koos nukleosiidfosfaatidega igasse kehanurka );
- täidab koensüümi funktsiooni (vitamiini- ja mineraalühendid, mis aitavad varustada rakke toitainetega);
- komplekssed ja tsüklilised mononukleotiidid on hormoonide bioloogilised juhid, mis levivad koos verevooluga ja suurendavad ka närviimpulsside mõju;
- reguleerivad allosteeriliselt pankrease kudede poolt toodetud seedeensüümide aktiivsust.
Nukleotiidid on osa nukleiinhapetest. Need on ühendatud kolme ja viie fosfodiestersidemega. Oma elu molekulaarbioloogiale pühendanud geneetikud ja teadlased jätkavad nukleosiidfosfaatide laboratoorseid uuringuid, nii et igal aastal saab maailm nukleotiidide omaduste kohta veelgi huvitavamat teada.
 Nukleotiidjärjestus on teatud tüüpi aminohapete paigutuse geneetiline tasakaal ja tasakaal DNA struktuuris, omapärane estrijääkide paigutuse järjekord nukleiinhapete koostises.
Nukleotiidjärjestus on teatud tüüpi aminohapete paigutuse geneetiline tasakaal ja tasakaal DNA struktuuris, omapärane estrijääkide paigutuse järjekord nukleiinhapete koostises.
Selle määramiseks kasutatakse traditsioonilist analüüsiks valitud bioloogilise materjali järjestamise meetodit.
T – tümiin;
A – adeniin;
G – guaniin;
C – tsütosiin;
R – GA adeniin kompleksis guaniini ja puriini alustega;
Y – TC pürimidiini ühendid;
K – ketorühma sisaldavad GT nukleotiidid;
M – aminorühma kuuluv AC;
S – võimas GC, mida eristab kolm vesinikuühendit;
W – AT on ebastabiilsed, moodustades ainult kaks vesiniksidet.
Nukleotiidide järjestus võib muutuda ja ladina tähtedega tähistamine on vajalik juhtudel, kui eetriühendite järjestus on teadmata, ebaoluline või esmaste uuringute tulemused on juba olemas.
Suurim hulk nukleosiidfosfaatide variante ja kombinatsioone on iseloomulik DNA-le. RNA eetriühendite kirjutamiseks piisab sümbolitest A, C, G, U. Viimase tähetähisena on aine uridiin, mida leidub ainult RNA-s. Sümboolsete märgete jada kirjutatakse alati ilma tühikuteta.
Kasulik video: nukleiinhapped (DNA ja RNA)
Mitu nukleotiidi on DNA-s
Selleks, et mõista võimalikult üksikasjalikult, millest me räägime, peaks teil olema selge arusaam DNA-st endast. See on eraldi tüüpi molekulid, millel on piklik kuju ja mis koosnevad struktuurielementidest, nimelt nukleosiidfosfaatidest. Mitu nukleotiidi on DNA-s? Seda tüüpi eetriühendeid, mis on osa DNA-st, on 4 tüüpi. Need on adeniin, tümiin, tsütosiin ja guaniin. Kõik need moodustavad ühe ahela, millest moodustub DNA molekulaarstruktuur.
DNA struktuuri dešifreerisid esmakordselt 1953. aastal Ameerika teadlased Francis Crick ja James Watson. Üks desoksüribonukleiinhappe molekul sisaldab kahte nukleosiidfosfaadi ahelat. Need on paigutatud nii, et need näevad välja nagu ümber oma telje keerduv spiraal.
Märge! Nukleotiidide arv DNA-s on konstantne ja piirdub vaid nelja tüübiga – see avastus on toonud inimkonna lähemale inimese täieliku geneetilise koodi dešifreerimisele.

Sel juhul on molekuli struktuuril üks oluline tunnus. Kõigil nukleotiidahelatel on komplementaarsuse omadus. See tähendab, et üksteise vastas asetatakse ainult teatud tüüpi eeterlikud ühendid. On teada, et adeniin asub alati tümiini vastas. Tsütosiini vastas ei leidu ühtegi teist ainet peale guaniini. Sellised nukleotiidipaarid moodustavad komplementaarsuse põhimõtte ja on lahutamatud.
Kaal ja pikkus
 Keeruliste matemaatiliste arvutuste ja laboratoorsete uuringute abil suutsid teadlased kindlaks teha desoksüribonukleiinhappe molekulaarstruktuuri moodustavate eetriühendite täpsed füüsikalised ja bioloogilised omadused.
Keeruliste matemaatiliste arvutuste ja laboratoorsete uuringute abil suutsid teadlased kindlaks teha desoksüribonukleiinhappe molekulaarstruktuuri moodustavate eetriühendite täpsed füüsikalised ja bioloogilised omadused.
On teada, et ühe polüpeptiidahela aminohapetest koosneva ühe rakusisese jäägi pikendatud pikkus on 3,5 ongströmi. Ühe molekulaarse jäägi keskmine mass on 110 amu.
Lisaks eraldatakse ka nukleotiidi tüüpi monomeere, mis ei moodustu mitte ainult aminohapetest, vaid sisaldavad ka eetrikomponente. Need on DNA ja RNA monomeerid. Nende lineaarset pikkust mõõdetakse otse nukleiinhappe sees ja see on vähemalt 3,4 ongströmi. Ühe nukleosiidfosfaadi molekulmass on 345 amu piires. Need on algandmed, mida praktikas kasutatakse laboritööd pühendatud eksperimentidele, geeniuuringutele ja muule teadustegevusele.
Meditsiinilised nimetused
Geneetika kui teadus arenes välja perioodil, mil inimeste ja teiste elusolendite DNA struktuuri ei uuritud. molekulaarne tase. Seetõttu nimetati premolekulaarse geneetika perioodil nukleotiidsidemeid DNA molekuli struktuuri väikseimaks elemendiks. Seda tüüpi olulisi aineid on nii varem kui ka praegu kasutatud. See võib olla spontaanne või indutseeritud, mistõttu kasutatakse terminit "recon" ka kahjustatud struktuuriga nukleosiidfosfaatide tähistamiseks.
Nukleotiidsidemete lämmastikuühendites võimaliku mutatsiooni esinemise mõiste määratlemiseks kasutatakse terminit "mouton". Need nimetused on laboritöös nõutavamad bioloogiline materjal. Neid kasutavad ka geeniteadlased, kes uurivad DNA molekulide ehitust, päriliku informatsiooni edastamise viise, selle kodeerimise meetodeid ning võimalikke geenikombinatsioone, mis on saadud kahe seksuaalpartneri geneetilise potentsiaali liitmise tulemusena.
Kokkupuutel
 Suur Tartari - Venemaa impeerium Inimkonna iidsest ajaloost
Suur Tartari - Venemaa impeerium Inimkonna iidsest ajaloost Vürst Ivan Vassiljevitš 1472
Vürst Ivan Vassiljevitš 1472 “A.P. Platonovi lugu “Tundmatu lill” (miniarvustus). Andrei Platonovi loost "Tundmatu lill" Mida tõeline muinasjutt Tundmatu lill mulle õpetas
“A.P. Platonovi lugu “Tundmatu lill” (miniarvustus). Andrei Platonovi loost "Tundmatu lill" Mida tõeline muinasjutt Tundmatu lill mulle õpetas