Muutuva valentsiga metalliioonide kelaatorid. Metalliioonide bioanorgaaniline keemia
Muutuva valentsiga metalliioonidel (Fe2+, Cu+, Mo3+ jne) on elusorganismides kahekordne roll: ühelt poolt on nad vajalikud kofaktorid tohutule hulgale ensüümidele, teisalt ohustavad nad raku eluiga, kuna nende olemasolu suurendab väga reaktiivsete hüdroksüül- ja alkoksüradikaalide moodustumist:
H202 + Me"n > OH' + OH" + Me(n+|)+
ROOOH + Men+ > 10* + OH" + Me(n+|>+.
Seetõttu kelaatühendid (kreeka keelest "kelaat" - "krabi küünis"), mis seovad muutuva valentsiga metalliioone (ferritiin, hemosideriin, transferriinid; tseruloplasmiin; piim- ja kusihape; mõned peptiidid) ja takistavad seeläbi nende osalemist lagunemises. peroksiidide reaktsioonid, mis on organismi antioksüdantse kaitse oluline komponent. Arvatakse, et kelaatorid on peamised seerumivalkude ja raku retseptorite kaitsmisel oksüdatsiooni eest, kuna rakkudevahelistes vedelikes rakumembraanidest hästi tungivate peroksiidide ensümaatiline lagunemine puudub või on oluliselt nõrgenenud. Muutuva valentsusega metalliioonide sekvestreerimise kõrget usaldusväärsust kelaativate ühendite abil tõendab Thomas W. O'Hallorani rühma (mudelina kasutati pärmirakke) paljastatud tõsiasi, et vabade* vaseoonide kontsentratsioon tsütoplasmas ei ületa 10"18 M - see on mitu suurusjärku vähem kui 1 Cu aatom raku kohta.
Lisaks kõrge ioonisidumisvõimega "professionaalsetele" kelaatijatele on olemas nn "oksüdatiivse stressi poolt aktiveeritud raua kelaatorid". Nende ühendite afiinsus raua suhtes on suhteliselt madal, kuid oksüdatiivse stressi tingimustes oksüdeeritakse need kohaspetsiifiliselt, mis muudab need tugeva rauda sidumisvõimega molekulideks. Arvatakse, et see lokaalne aktiveerimisprotsess minimeerib "tugevate kelaatijate" potentsiaalset toksilisust kehas, mis võib häirida raua metabolismi. Mõned kelaatorid, nagu metallotioneiinid, seovad imetajatel raskmetallide aatomeid (Chn, Sb, Sh,...) ja osalevad nende detoksikatsioonis.
Veel teemal MUUTUVVALENTSUSEGA METALLIIOONIDE KEELAATID:
- NovikA. A., Ionova T.I.. Meditsiini elukvaliteedi uurimise juhend. 2. trükk / Toim. akad. RAMS Y.L. Ševtšenko, - M.: JSC "OLMA Media Group" 2007, 2007
- 3. PEATÜKK KESKMISE JA KÕRGSAGEDUSEGA VAHDUVA VOOLU RAVIKASUTAMINE
- Test kehaasendi muutmisega (ortostaatiline test)
- Raskmetallide soolade farmakoloogilise toime spekter
Pärast selle peatüki uurimist peaks õpilane:
tea
Leelis- ja leelismuldmetalliioonide ökoloogilised ja füsioloogilised põhiandmed, plii mõju inimorganismile, raskmetallide aatomite migratsiooni vormid atmosfääris ja hüdrosfääris;
suutma
Tee kindlaks vee sobivus erinevateks otstarveteks;
oma
- kaitsemeetodid toksiliste metalliioonide inimtekkeliste mõjude eest.
Sõltuvalt nende käitumisest elussüsteemides jagatakse ained, sealhulgas metalliioonid, viide tüüpi: organismile vajalik; stimulandid; inertne, kahjutu; raviained; mürgine. Organismile peetakse vajalikuks ainet, mille defitsiit põhjustab organismis funktsionaalseid häireid, mida saab kõrvaldada selle aine sisseviimisega. Vajadus on organismispetsiifiline omadus ja seda tuleks eristada stimulatsioonist. On palju näiteid, kui nagu stimulandid Ilmuvad nii olulised kui ka mitteolulised metalliioonid. Mõned metallid ja metalliioonid teatud kontsentratsioonides on inertne, kahjutu ja ei avalda kehale mingit mõju. Seetõttu kasutatakse kirurgiliste implantaatidena sageli inertseid metalle - Ta, Pt, Ag, Au. Paljud metalliioonid võivad teenida raviained;
Joonisel fig. 6.1 annab aimu kehakudede bioloogilisest reaktsioonist piisavas koguses, näiteks kerjusest, tarnitavate metalliioonide kontsentratsiooni suurenemisele.
Riis. 6.1. Bioloogiline reaktsioon olenevalt nõutavast kontsentratsioonist(tugev kõver)ja ohtlik(katkendlik kõver)ained
(kahe kõvera suhteline asukoht kontsentratsiooni skaala suhtes on suvaline)
Tahke kõver näitab kohest positiivset reaktsiooni suureneva kontsentratsiooniga, alustades nullist (eeldatakse, et sissetulev vajalik aine küllastab oma seondumiskohad ega astu muudesse interaktsioonidesse, mis on tegelikult täiesti võimalikud). See tahke kõver kirjeldab optimaalset taset, mis hõlmab paljude metalliioonide laia kontsentratsioonivahemikku. Metalliioonide kontsentratsiooni suurenemise positiivne mõju läbib maksimumi ja hakkab langema negatiivsete väärtusteni: keha bioloogiline reaktsioon muutub negatiivseks ja metall muutub mürgiseks aineks.
Katkendlik kõver joonisel fig. 6.1 näitab organismi bioloogilist reaktsiooni täiesti mürgisele ainele, millel ei ole vajaliku või stimuleeriva aine toimet. See kõver järgib teatud mahajäämust, mis näitab, et elusorganism suudab "leppida" väikese koguse mürgise ainega (lävikontsentratsioon), kuni selle toksiline toime muutub domineerivaks.
Joonisel fig. 6.1 esitab muidugi teatud üldistava pildi; Igal ainel on oma spetsiifiline kõver "bioloogiline reaktsioon - kontsentratsioon" koordinaatides. Jooniselt järeldub ka, et olulised ained võivad liigsel tarbimisel muutuda isegi mürgiseks. Peaaegu iga üleliigne aine muutub paratamatult ohtlikuks (isegi kui see mõju on kaudne), näiteks teiste vajalike ainete piiratud imendumise tõttu. Looma keha hoiab ainete kontsentratsiooni optimaalses vahemikus füsioloogiliste protsesside kompleksi nn homöostaas. Kõigi oluliste metalliioonide kontsentratsioon eranditult on homöostaasi range kontrolli all; Paljude metalliioonide homöostaasi üksikasjalik mehhanism jääb praeguste uuringute valdkonda.
Inimorganismile (ja loomadele) vajalike metalliioonide loetelu on toodud tabelis. 6.1. Kuna uuringud jätkuvad ja katsemeetodid muutuvad rafineeritumaks, peetakse mõnda varem mürgiseks peetud metalli nüüd hädavajalikuks. Tõsi, Ni 2+ inimorganismile vajalikkust pole veel tõestatud. Eeldatakse, et ka teisi metalle, näiteks tina, võib pidada imetajate jaoks hädavajalikuks. Tabeli teine veerg. 6.1 näitab vormi, milles antud metalliioon eksisteerib pH = 7 juures ja võib esineda vereplasmas, kuni ühineb teiste ligandidega. Tahkel kujul FeO(OH) ja CuO plasmas ei leidu, kuna nii Fe 3+ kui ka Cu 2+ moodustavad komplekse valgu makromolekulidega. Tabeli kolmandas veerus. Tabelis 6.1 on näidatud iga täiskasvanud inimese kehas tavaliselt esineva olulise elemendi tüüpiline üldkogus. Vastavalt sellele on metalliioonide kontsentratsioonid plasmas toodud neljandas veerus. Ja viimane veerg soovitab iga vajaliku metalliiooni päevase tarbimise kogust, kuid need soovitused võivad muutuda.
Tabel 6.1
Olulised metalliioonid
|
Kuju pi I = 7 juures |
Kontsentratsioon vereplasmas, mmol |
Päevane tarbimine, g |
||
Vastuseks välisele sekkumisele on elusorganismil teatud detoksikatsioonimehhanismid, mis piiravad või isegi kõrvaldavad mürgise aine. Spetsiifiliste metalliioonidega seotud võõrutusmehhanismide uurimine on varajases staadiumis. Paljud metallid muudetakse organismis vähemkahjulikeks vormideks järgmistel viisidel: lahustumatute komplekside moodustumine soolestikus; metalli transport veres teistesse kudedesse, kus seda saab immobiliseerida (näiteks Pb 2+ luudes); muundamine maksa ja neerude poolt vähem toksilisemaks või vabamaks vormiks. Seega suurendavad inimese maks ja neerud vastusena toksiliste ioonide Cd 2+, Hg 2+, Pb 2+ jt toimele metallotioneiinide – madala molekulmassiga valkude – sünteesi, milles ligikaudu 10 (61-st) aminohappejäägid on tsüsteiin. Sulfhüdrüül-SH-rpynni kõrge sisaldus ja hea vastastikune paigutus annavad võimaluse metalliioonide tugevaks sidumiseks.
Metalliioonide mürgisteks muutumise mehhanisme on üldiselt lihtne ette kujutada, kuid ühegi konkreetse metalli puhul on raske täpselt kindlaks teha. Metalliioonid stabiliseerivad ja aktiveerivad paljusid valke; Ilmselt vajavad kõik ensüümid U 3 toimimiseks metalliioone. Konkurentsi oluliste ja toksiliste metalliioonide vahel valkude seondumiskohtade pärast pole raske ette kujutada. Paljudel valgu makromolekulidel on vabad sulfhüdrüülrühmad, mis võivad interakteeruda toksiliste metalliioonidega nagu Cd 2+, Hg 2+, Pb 2+; Laialdaselt arvatakse, et see reaktsioon on viis loetletud metalliioonide toksilisuse avaldumiseks.
Samas pole täpselt kindlaks tehtud, millised valgu makromolekulid põhjustavad elusorganismile kõige tõsisemaid kahjustusi. Mürgised metalliioonid on jaotunud paljude kudede vahel ja pole mingit garantiid, et suurimad kahjustused tekivad seal, kus metalliiooni on kõige rohkem. Seda on näidatud näiteks Pb 2+ ioonide puhul: olles enam kui 90% (nende kogusest kehas) luudes immobiliseeritud, jäävad need mürgiseks, kuna 10% jaotub teistesse keha kudedesse. Tõepoolest, Pb 2+ ioonide immobiliseerimist luudes võib pidada võõrutusmehhanismiks. Seda tüüpi toksilisust, mis on põhjustatud geneetilistest haigustest (nt Cooley aneemia, millega kaasneb liigne rauasisaldus), selles peatükis ei käsitleta.
Meie ülevaade ei puuduta metalliioonide võimalikku kantserogeenset aktiivsust. Captseroheppost - see on kompleksne nähtus, mis sõltub looma tüübist, elundist ja tema arengutasemest, sünergiast teiste ainetega. Metalliioonid ja nende kompleksid võivad samuti toimida vähivastased ained. Metalliiooni mürgisus ei ole tavaliselt seotud selle vajadusega kehale. Siiski on toksilisusel ja vajalikkusel üks ühine joon: üldiselt on metalliioonide vastastikune sõltuvus nii üksteisest kui ka metalli- ja mittemetalliioonide üldise panuse osas nende tõhususse. Oluliste metalliioonide kättesaadavus sõltub nende koostoimest tarbitava toiduga; Lihtne toitumise adekvaatsus seda olukorda ei rahulda. Näiteks juurviljadest saadav raud imendub halvasti neis sisalduvate kompleksi moodustavate ligandide tõttu ning Zn 2+ ioonide liig võib pärssida Cu 2+ imendumist. Samamoodi on Cd 2+ toksilisus rohkem väljendunud Zn 2+ vaegusega süsteemis ja Pb 2+ toksilisust süvendab Ca 2+ defitsiit. Selline antagonism ja vastastikune sõltuvus raskendab oluliselt katseid leida ja selgitada vajalikkuse ja toksilisuse põhjuseid.
Paljude metalliioonide puhul tekib äge mürgisus siis, kui tekib äkiline "löök" suure metalliannusega; sel juhul ilmnevad muud mõjud ja sümptomid kui kroonilise mürgistuse korral; Krooniline mürgistus tekib metalli väikeste annuste saamisel, kuid pika aja jooksul.
Metalliioonide kõige tõsisemad toksilised mõjud tekivad tolmu sissehingamisel, mis esineb tavaliselt tööstuslikes tingimustes. Eriti ohtlikud on osakesed läbimõõduga 0,1 - 1 mikronit, mis adsorbeeritakse tõhusalt kopsudesse. Pange tähele, et kopsud neelavad metalliioone, mis seejärel sisenevad kehavedelikesse, kümme korda tõhusamalt kui seedetrakt. Näiteks radioaktiivse nlutoonium-239 (mis eraldab aktiivseid alfaosakesi poolestusajaga 24,4 tuhat aastat) suurim oht tuleneb mitte plutooniumi imendumisest toidus, vaid plutooniumipulbri adsorptsioonist kopsukoes.
Lenduvad metalliühendid, nagu elavhõbeda, plii ja tina karbonüül- ja alküülühendid, imenduvad kopsudesse kergesti ja võivad põhjustada ägedat metallimürgitust. Siit järeldus: igasugust metalliioonide sissehingamist tuleks vältida!
Leelismetalli ioonid. Ükski leelismetallidest pole eriti mürgine. Homöostaas hoiab mõlema oluliste ioonide Na + ja K + kontsentratsiooni (vt tabel 6.1) normaalsel füsioloogilisel tasemel. Mõlema elemendi roll on seedimisel oluline. Lisaks spetsiifilistele toimetele mängivad need metalliioonid elusorganismides kaht olulist rolli: nad määravad osmootse tasakaalu mõlemal pool membraani ja pakuvad positiivseid vastasioone anioonidele, nagu HPO, HCO3 ja orgaanilised molekulid, millest paljud on anioonid. Seega on peamised rakkudevahelised ja intratsellulaarsed vastasioonid vastavalt Na + ja K +.
Teised leelismetalliioonid võivad mõnes füsioloogilises protsessis konkureerida Na + ja K + ioonidega. Inimkehas sisaldab rakusisene vedelik koos K1 ioonidega ligikaudu 0,3 g Rb +. Väikestes kogustes võib esineda ka Cs+; märkimisväärne kogus 37 Cs (T| 2 = 30 aastat) ilmneb ainult radioaktiivse kiiritamise korral. Suurim sisemistest allikatest pärit sugunäärmete radioaktiivsuse doos on tavaliselt 20 mrem aastas ja see saadakse looduslikust kaaliumist, mida rakusiseses vedelikus tingimata leidub.
Liitium. Li* on kasutatud maniakaal-depressiivse psühhoosi raviks üle 50 aasta; Ühendkuningriigis saab seda ravimina keskmiselt iga kaks tuhat inimest. Li 2 C0 3 suukaudne tarbimine tõstab liitiumi kontsentratsiooni vereplasmas 1 mM-ni, mis silub märkimisväärselt paljude patsientide meeleolumuutusi. Kuid terapeutilise toime saavutamiseks vajalik metallitase võib paraku avaldada toksilist mõju, nagu neerufunktsiooni pärssimine ja kesknärvisüsteemi häired. Liitiumioonide toime olemus pole ikka veel selge; see võib muuta rakusisest interaktsiooni. Li+ mõjutab paljusid ensüüme, sealhulgas neid, mis osalevad glükolüüsis. Paljud biokeemikud usuvad, et Li + asendab Na b või K + ioone, kuid nende maht on vastavalt kolm või kuus korda suurem kui liitium. Seetõttu peaks selline asendus valgu makromolekulides põhjustama muutuse vastavate metalliõõnsuste struktuuris; teisalt on Li + ioon veidi suurem kui Mg 2+ ioon. Liitium moodustab tavaliselt tugevamaid komplekse kui Na + ja K +, kuid palju nõrgemaid kui Mg 2+. Psühhoosi ravis kasutatakse liitiumi ja magneesiumi võrreldavates kontsentratsioonides ning Li + hõivab need seondumiskohad, mida Mg 2+ ei hõivata; kui kõik võimalikud kohad on hõivatud magneesiumiga, tõrjub Li* välja Na + ja K +. Kõik need leelismetalliioonid astuvad vahetusreaktsioonidesse rohkem kui 10 3 korda kiiremini kui Mg 2+ ioon. Just see tegur võib seletada Mg-d sisaldavate ensüümide aktiivsuse muutust liitiumi sisseviimisel.
Magneesium. See metall Mg 2+ iooni kujul on vajalik nii taime- kui loomaorganismidele. Taimedes on Mg 2+ kelaaditud nelja lämmastikuaatomiga klorofülli tsüklilise struktuuri pürroolitsüklites, harv juhus magneesiumi koordinatsioonist lämmastikuga. Loomorganismides on Mg 2+ vajalik kofaktor igas reaktsioonis, mis hõlmab adenosiintrifosfaati (ATP). Samuti mängib see vastasiooni rolli DNA kaksikheeliksi stabiliseerimisel, mille igas ahela lülis on negatiivselt laetud fosfaatrühmad. Magneesiumioonide olemasolu suurendab linkide õige sidumise tõenäosust. Nukleosiidfosfaatidega nagu ATP koordineerituna seondub Mg2+ ainult fosfaatrühmadega. Mg 2+ ioonid on hädavajalikud neuromuskulaarseks ülekandeks ja lihaste kontraktsiooniks. Püsiv homöostaas hoiab praktiliselt tervetel inimestel Mg 2+ taseme vereplasmas 0,9 mM tasemel. Mg 2+ puudust esineb palju sagedamini ja alkoholismi puhul tundub see olevat kohustuslik olukord. Kuna tõsine magneesiumipuudus on üsna haruldane nähtus, on sümptomite kohta vähe andmeid. Selle sümptomiteks on deliirium tremens ja neuromuskulaarsed ilmingud, sealhulgas külmavärinad, krambid, jäsemete tuimus ja värinad. Madal Mg 2+ tase võib põhjustada hüpokaltseemiat, mille puhul metaboolselt labiilset mineraali ei saa luudest mobiliseerida. Nii Mg 2+ kui Ca 2+ taset kontrollib paratüreoidhormoon negatiivse tagasiside mehhanismi kaudu. Magneesium on üsna kergelt mürgine. Suures koguses Mg 2+ soolade võtmine põhjustab oksendamist. Neerupuudulikkusega patsientidel, kes said magneesiumi hapet neutraliseerivates ravimites, võivad tekkida pikaajalised toksilisuse sümptomid. Viimased võivad mõjutada kesknärvisüsteemi, hingamiselundeid ja kardiovaskulaarsüsteemi.
Kaltsium. Kaks leeliselist iooni Na~ ja K+ ning kaks leelismuldmetalli iooni Mg 2+ ja Ca 2+ moodustavad kokku enam kui 99% metalliioonide kogusest inimkehas. Kaltsiumi Ca 2+ kujul leidub kehas rohkem kui teisi metalliioone. Rohkem kui 99% sellest on osa luudest ja hambaemailist hüdroksoapatiidi Ca 5 (P0 4) 3 (0H) kujul. Kaltsium mängib lahuses olulist rolli paljudes protsessides, sealhulgas lihaste kokkutõmbumises, vere hüübimises, närviimpulssides, mikrotuubulite moodustumisel, rakkudevahelises suhtluses, hormonaalsetes reaktsioonides, eksotsütoos, viljastumine, mineralisatsioon, aga ka rakkude ühinemine, adhesioon ja kasv. Paljud kaltsiumiooni loetletud aktiivsused on seotud interaktsioonidega valgu makromolekulidega, mida Ca 2+ ioon suudab stabiliseerida, aktiveerida ja moduleerida. Kõik seni valkudes Ca 2+ ioonide seondumiskohad koosnevad hapnikuaatomitest. Ca 2+ kontsentratsioonigradient rakkudevahelistes ja intratsellulaarsetes vedelikes ületab oluliselt ülejäänud kolme bioloogiliselt olulise leelis- ja leelismuldmetalliiooni (Na +, K, Mg 2+) gradiente Ca 2+ vaba kontsentratsioon rakkudevahelistes vedelikes on umbes 1,3 mM, samas kui paljudes intratsellulaarsetes vedelikes on see silmatorkavalt madal (0,1 µM või isegi madalam 20 000-kordse kontsentratsioonigradiendi korral). Stimuleerimisel võib madal rakusisene kontsentratsioon tõusta 10 korda, millega kaasnevad valgu konformatsioonilised muutused. makromolekulid, mille dissotsiatsioonikonstant on mikromoolides. Mõnede rakusiseste valkude konformatsiooniline tundlikkus kaltsiumi kontsentratsiooni muutuste suhtes mikromolaarsel tasemel on viinud arusaamiseni Ca 2+ rollist teist tüüpi intratsellulaarse vahendajana. Soovitatav päevane annus ( 800 mg) Ca 2+ on võimalik saada liitri piima joomisega – ainsa kaltsiumirikka allikaga. Kaltsiumipuudus väljendub kasvupeetuses, halbades hammastes ja muudes vähemnähtavates defektides. Üheks selliseks varjatud defektiks on soovimatute või toksiliste metalliioonide suurenenud neeldumine Ca 2+ -puudulikus süsteemis. Homöostaasi mehhanism, mis kontrollib imendumist soolestikust, kontrollib Ca 2+ taset inimestel. Kaltsiumi peetakse mittetoksiliseks. Luu mineraalide ladestumist pehmetesse kudedesse ei põhjusta liigsed Ca 2+ ioonid, vaid D-vitamiini kõrgenenud tase. Samas võib kõrge Ca 2+ sisaldus toidus pärssida teiste organismile vajalike metallide imendumist soolestikust.
Baarium ja strontsium. Ba 2+ on mürgine oma antagonismi tõttu K+-ga (kuid mitte Ca 2+-ga). See seos on selge näide Ba 2+ ja K + ioonraadiuste sarnasuse suuremast tähtsusest kui laengu identsus (kahel leelismuldmetalli ioonil Ba 2+ ja Ca 2+ on erinevad raadiused). Baariumioon on lihasmürk, siinne ravi seisneb K + soolade intravenoosses manustamises. Kuigi Ba 2+ ioonid on veel soolestikus, võttes lahustuvaid sooli SO| _ põhjustab lahustumatu baariumsulfaadi moodustumist, mis ei imendu. BaSO| kasutatakse seedetrakti uuringutes radioaktiivse materjalina. Inimkeha sisaldab luudes ligikaudu 0,3 g Sr 2+. See summa ei kujuta endast mingit ohtu; kuid strontsium on viimastel aastatel radioaktiivse sademe tõttu ulatuslikult saastunud 90 Sr kujul (G 1/2 = 28 aastat).
Berüllium. Be 2+ moodustab happelises keskkonnas lahustumatu hüdroksiidi Be(OH) 2, mis vähendab imendumist soolestikust. Berülliumi sisaldava tolmu sissehingamine põhjustab kroonilist pulmonaalset granulomatoosi (nimetatakse berülliumiks) või kahjustusi kopsudes; haigus areneb aeglaselt ja on sageli surmav. Luminofoorlampe tootvate tehaste töötajad, kus berülliumoksiidi kasutatakse fosforestseeruva ainena, on langenud berülliumhaiguse ohvriks. (Selline tootmine on juba peatatud.) Juba miljondik kehamassist berülliumi annus on surmav. Be 2+ ringleb kehas kolloidfosfaadi kujul ja sulandub järk-järgult luuskeletti. Hüdroksiidi ja fosfaadi komplekside moodustumine toimub vastavalt ülaltoodud põhimõtetele (väikese suurusega, kuid suure laengutihedusega kahevalentsete ioonide puhul). Be 2 ~ inhibeerib paljusid fosfataasi tüüpi ensüüme ja on tugevaim teadaolev aluselise fosfataasi inhibiitor. Berüllium pärsib ka magneesiumi ja kaaliumi poolt aktiveeritud ensüüme ning häirib DNA replikatsiooni. On näidatud, et "kelaatravi" (kelaativate ainete, näiteks etüleendiamiintetraäädikhappe võtmine) on kroonilise berülliumimürgistuse all kannatavate inimeste kehast Be 2+ eemaldamisel ebatõhus. On ilmselge, et sellise latentse (pikaajalise) mürgisusega ohtliku ainega nagu berüllium tuleks suhtuda väga ettevaatlikult ja parem on see üldse ringlusest eemaldada.
Lantaniidid. Lantaniidid sisaldavad 15 elementi, alates lantaanist aatomnumbriga 57 kuni luteetiumini aatomnumbriga 71. Kõiki neid leidub bioloogilistes süsteemides ainult +3 oksüdatsiooniastmes. Gadoliiniumi Gd 3+ puhul – selle seeria keskmine liige (aatomnumber 64) – ühtib ioonraadius täpselt Ca 2+ ioonraadiusega. Kuna aatomi suuruse sarnasus on olulisem kui laengu võrdsus, asendavad lantaniidid kaltsiumi paljudes bioloogilistes süsteemides. See lantaniidi asendus ei ole oluline, kui metalliioonil on valdavalt struktuurne roll, kuid sellel võib olla inhibeeriv või aktiveeriv toime, kui metalliioon on aktiivses kohas. Lantaniidiioone kasutati väga laialdaselt Ca 2+ ioonide seondumiskohtade määramisel valgu makromolekulides. Ükski lantaniidi element ei ole bioloogiliselt oluline. Taimed peavad vastu lantaniidide kuhjumisele, takistades seeläbi lantaniidide ülekandumist inimestele peamiselt toiduahela kaudu. Lantaniidid on vesiioonina (3+) kuni pH = 6-ni, mil algab hüdroksokomplekside ja setete moodustumine. Nende fosfaadid on samuti lahustumatud. Selle tulemusena moodustavad lantaniidid soolestikus lahustumatud kompleksid ja imenduvad seetõttu halvasti. Ühtegi neist ei peeta mürgiseks.
Alumiiniumist. Alumiiniumi, mis on maakoores kõige levinum metall, leidub elusorganismides harva, arvatavasti seetõttu, et seda on keeruline saada osana keerukatest maavaradest. Tavaliselt sisaldab täiskasvanud inimese keha 61 mg alumiiniumi, millest suurem osa on sissehingamise tulemusena kopsudes. Ainus alumiiniumkatioon A1 3+ neutraalsetes lahustes moodustab lahustumatu hüdroksiidi A1(OH) 3 ja selle alusel tugevalt ristseotud hüdrokso- ja oksoühendeid. Just selliste osakeste ja lahustumatu A1P0 4 moodustumine piirab A1 3+ imendumist seedetraktis. Pärast imendumist on alumiiniumi suurim kontsentratsioon ajus. Neerufunktsiooni halvenemine vähendab oluliselt organismi võimet eritada A1 3+. Alumiiniumi kõrge sisaldus põhjustab AlP0 4 moodustumise tõttu fosfaadi vähenemist. Seda metalli on vees ja toidus võimalik vaid vähesel määral ning sellistes kontsentratsioonides pole A1 3+ üldsegi eriti mürgine. Al 3+ (nagu ka Hg 2+ ja Pb 2+) sattumine happevihmadega linnade veevärki toob kaasa suurema metallisisalduse, mis on juba muutumas probleemiks. Vette sattuvad metalliioonid võivad kaladele kujutada endast palju tõsisemat ohtu kui happesus. Piiratud kogused Ca 2+ ja Mg 2+ näivad suurendavat alumiiniumi potentsiaalset toksilisust. A1 3+ toksiline toime avaldub kõhukinnisuse ja närvisüsteemi häiretena. Alumiiniumi suurenenud kontsentratsioon ajus on seotud Alzheimeri tõve, selliste häiretega nagu dementsus ja isegi surmaga, peamiselt vanematel inimestel. Kaasaegse meditsiini kontseptsioonide kohaselt ei ole alumiinium aga tõenäoliselt haiguse peamine põhjus, vaid koguneb niigi ebatervislikku ajju või toimib ühena paljudest teguritest. Igatahes on väga murettekitav märk asjaolu, et vanem põlvkond kasutab alumiiniumi sisaldavaid higistamisvastaseid aineid ja võtab sisse ka suures koguses antatsiide (happesust neutraliseerivad ravimid). Patsientidel, keda dialüüsitakse suure A1 3+ kontsentratsiooniga vees, võib tekkida "dialüüsidementsus".
Kroom. Kroom on traditsiooniliselt lisatud vajalike mikroelementide loendisse. Inimkeha sisaldab umbes 6 mg kroomi, mis on jaotunud paljude kudede vahel. Kuigi vajalikke annuseid ei ole kindlaks määratud, peaksid need olema väga väikesed. Vajalikku kroomi taset on keemiliste või biokeemiliste meetodite abil raske hinnata. Samuti jääb teadmata kroomi vajaduse põhjus. Kuigi 25 aastat on möödunud ajast, mil Cr 3+ esmakordselt pakuti välja glükoositaluvuse faktori komponendiks, jääb kompleksi enda olemus teadmata ja mõned sellise kompleksi jaoks välja pakutud struktuurid näivad olevat alusetud. pH = 7 juures on kõige levinum ühend Cr(OH)2, kuid selle inertsel polünukleaarsel komplekssel kujul. Isegi kroom(III)heksaakvaaviooni kujul kulub veemolekuli vahetus lahustiga mitu päeva. Just see inertsus piirab ilmselt Cr(III) rolli ainult struktuursete funktsioonidega. Kui kroom osaleb sellegipoolest kiiretes reaktsioonides, siis toimib see neis Cr(II)na. Suhkrud võivad toimida kroomi potentsiaalsete ligandidena. Glükoos on selle metalli sidumiseks suhteliselt halb ligand, kuid see piirang ei pruugi mõnes kolmevalentses kroomikompleksis rolli mängida. Kolmevalentne Cr(III) on üks kõige vähem toksilisemaid metalliioone; tugev oksüdeerija kuuevalentne Cr (VI) on juba mürgisem. pH juures
Molübdeen. See metall esineb tavaliselt Mo(VI) kujul ja molübdaat MoO|“ adsorbeerub seedetraktis. Molübdeen esineb taimedes ensüümi lämmastiku kofaktorina. Ksantiinoksüdaasil (mis katalüüsib kusihappe moodustumist loomadel) on adeniini dinukleotiidi kofaktorite osana kaks Mo-aatomit, kaheksa Fe-aatomit ja kaks flaviinitsüklit. Molübdeeni toksilisus on vase või väävli mürgisuse tasemel. Mäletsejalistel, keda toidetakse molübdeeniga rikastatud ja vasest vaesestatud söödaga, tekivad kasvajad, millega kaasneb kasvu aeglustumine, aneemia ja luuhaigused. Inimestel põhjustab sarnase molübdeeni ja vase suhtega dieet podagra sümptomeid. Vasepreparaatide võtmine on kasulik molübdeenimürgistuse saanud loomadele. Eriti mürgiseks metalliks ei peeta ei molübdeeni ega sellega seotud volframi, mis pole organismile hädavajalik ja pärsib ksantiinoksüdaasi aktiivsust.
Mangaan. Mangaani puhul on teada mitu oksüdatsiooniastet, kuid on tõendeid selle kohta, et see metall ei osale redoksreaktsioonides ja oluline on ainult Mn 2+; Mn 3+ on veeioonina ebastabiilne pH > 0 juures ja kui see ei ole komplekssel kujul, taandub neutraalsetes lahustes kergesti Mn 2+-ks. Puuduvad andmed selle kohta, mida põhjustab mangaani puudus inimkehas. Loomadel põhjustab selle puudus luude kasvu halvenemist, produktiivse funktsiooni langust ja võib-olla ka kolesterooli sünteesi pärssimist. Mangaan võib olla ensüümide kofaktor. Kuigi Mn 2+ aktiveerib paljusid ensüüme, on see aktiveerimine spetsiifiline, kuna sellel eesmärgil on tõhusad ka teised metalliioonid, näiteks Mg 2+. Mn 2+ kontsentratsioon vereplasmas on vaid üks tuhandik Mg 2+ kontsentratsioonist. Mangaan on peaaegu mittetoksiline, eriti Mn 2+ iooni kujul. Permanganaadi ioon MnOj on oma oksüdeerivate omaduste tõttu mürgine. Kõige tavalisem mangaanimürgistus tekib selle oksiidi sissehingamise tõttu tööstuslikus tootmises. Seda tüüpi krooniline toime võib põhjustada manganismi, mis põhjustab tõsist pöördumatut kesknärvisüsteemi ja aju kahjustust. Ilmselt avaldab liigne mangaan organismis mõju aju ensüümsüsteemidele. Kahjuks pole universaalseid tõhusaid vastumürke, vaid püütakse lihtsalt algpõhjus kõrvaldada.
Raud. Rauasisaldus inimkehas on 4 g, millest umbes 70%, s.o. 3 g leidub punastes verelibledes hemoglobiini kujul, ülejäänud osa rauavalkudes ja väike kogus mõnedes ensüümides. Soovitatavast ööpäevasest rauavajadusest 10-20 mg imendub vaid 10-20%, veidi suurem kogus hea homöostaasiga rauapuudusega inimestel. Raua imendumist pärsib lahustumatute hüdroksiidide, fosfaatide ja rasvhapetega komplekside moodustumine; seda soodustavad lahustuv suhkur ja askorbiinhappe kelaadid. Peaaegu kogu hemoglobiini lagunemisel iga päev vabanev 25 mg rauda suunatakse maksas tõhusalt ümber, nii et raua eluiga inimkehas ületab 10 aastat. Seetõttu piisab inimesele imendumisest alla 1 mg päevas (erandiks on menstruatsiooniperiood, mille jooksul naine kaotab umbes 20 mg rauda). Kõige levinum inimeste puudus maailmas on rauapuudus, mis mõjutab kuni 10% tööstuspiirkondades elavatest menopausieelsetest naistest; mõnes rühmas tõuseb see näitaja 100%-ni. Rauapuudus põhjustab aneemiat. Raud imendub Fe(II) kujul ja oksüdeerub veres Fe(III)-ks. Kuna Fe 3+ moodustab isegi happelistes vesilahustes täiesti lahustumatuid sademeid, kannab proteiin transferriin Fe 3+ verre. Kui transferriini Fe 3+ kandevõime on ammendatud, ladestub Fe(OH) 3 verre. Raua mürgisus mõjutab konkreetseid rühmi: USA-s sureb tuhandest lapsest umbes 10 aastas emadele valmistatud FeS0 4 mineraaltableti neelamise tõttu; kus toiduvalmistamine toimub raudpottides; raske maksapuudulikkuse all kannatavate alkohoolikute seas. Raua toksilisus on seotud seedetrakti haiguste, šoki ja maksakahjustusega.
Koobalt tuntud B12-vitamiini olulise komponendina, kelaaditud nelja ühendatud pürroolitsükliga kompleksseks korriini makrotsükliks. Inimese päevane vajadus B12-vitamiini järele on vaid 3 mikrogrammi ning selle puudus põhjustab aneemiat ja kasvupeetust. Teada on mitmeid B12-vitamiini vorme, mis toimivad ensüümide kofaktoritena metüülrühma ülekandereaktsioonis, aga ka muudes reaktsioonides, kus koobalti oksüdatsiooniaste muutub. Koobaltit leidub bioloogilistes süsteemides Co 2+ iooni kujul, ilma et see oleks seotud vitamiini B 12 -korrinoidtsükliga. See ioon on võimeline siduma nelja, viit ja isegi kuut doonoraatomit erinevat tüüpi koordinatsioonipolüeedrites. Sarnane võime on ka Zn 2+-l. Nendel kahel ioonil on kõigi koordinatsiooniarvude jaoks samad efektiivsed ioonraadiused, samuti üsna võrreldavad stabiilsuskonstandid. Paljude ligandidega kompleksides asendab Co 2+ mõnes ensüümis Zn 2+, andes sageli ka aktiivseid ensüüme. Kuna sellel on paardumata ^/- elektronid, kasutavad mõned spektrimeetodid tsinki sisaldavates valkudes spektriliselt inaktiivse tsingi omaduste uurimiseks Co 2+. Liigne Co 2+ stimuleerib luuüdi tootma punaseid vereliblesid; see vähendab ka kilpnäärme võimet joodi akumuleerida, s.t. Struuma võib olla aneemia koobaltisoolade võtmise tagajärg. Koobalt on näidanud kardiotoksilisust mõne innuka õllejooja jaoks, kes tarbivad rohkem kui kolm liitrit päevas. (Mõnes riigis lisatakse õllele vahu stabiliseerimiseks 10–4% koobaltisoolasid, et neutraliseerida jääkpesuvahendite mõju.) Kuigi ohvrite arv oli väiksem kui Co2+ aneemiaravimite puhul, on siiski selge, et etüül alkohol suurendab organismi tundlikkust koobaltimürgistuse suhtes ning pudeliõlles sisalduv SO 2 hävitab tiamiini (selle vitamiini puudus süvendab Co 2+ põhjustatud kardiotoksilisust).
Nikkel. Bioloogilistes süsteemides esineb nikkel peaaegu eranditult Ni(II) kujul. Kuigi nikli puhul on teatud tingimustel võimalik oksüdatsiooniaste +3, on selle esinemine kõrgelt arenenud organismides ebatõenäoline. Inimkeha sisaldab umbes 10 mg Ni 2+ ja selle tase vereplasmas on üsna kitsastes piirides, mis viitab homöostaasile ja võib-olla ka nikli vajadusele. Madal Ni2* tase stimuleerib loomi. See toimib taimeensüümi ureaasi kofaktorina. Ni 2 * aktiveerib koos teiste metalliioonidega loomade kehas teatud ensüüme, kuid selle vajalikkust inimesele pole veel tõestatud. Ni 2+ ioon on veel üks näide metallist, mis on suhteliselt mittetoksiline. Ja veel, tööstusaurud, eriti need, mis sisaldavad nikkelkarbonüül-Ni(CO) 4 (milles nikkel on formaalselt nullvalentses olekus), imenduvad kopsudesse kergesti ja on väga mürgised. Allaneelamisel põhjustab Ni 2+ ioon ägedat ebamugavustunnet seedetraktis. Krooniline nikli mürgistus põhjustab südame ja teiste kudede hävimist. Nikli mürgisuse põhjused on meile teadmata; see blokeerib ensüüme ja reageerib nukleiinhapetega.
Vask. Vase kontsentratsiooni organismis reguleerib homöostaas ja selle optimaalsed kontsentratsioonid on väga erinevad. Seetõttu pole vasepuudus ega selle mürgisus tavalised juhtumid. Vask on oluline kofaktor mitmete ensüümide jaoks, mis katalüüsivad mitmesuguseid redoksreaktsioone. Selle puudus põhjustab aneemiat, luu- ja sidekoe halba seisundit ning juuste pigmentatsiooni kadu. Võimalik, et Zn 2+ võtmine näiteks pillides võib tekitada vasepuuduse. Vask mõlemas valentsolekus, Cu(I) ja Cu(II), seondub hästi sulfhüdrüülrühmaga glutatioonis ja väävlit sisaldavates valkudes. Cu(II) oksüdeerib kaitsmata sulfhüdrüülrühma disulfiidrühmaks, redutseerub ise Cu(I)-ks, seega peab keha enne sulfhüdrüülrühma oksüdeerumist Cu(I) siduma. Umbes 95% vereplasmas leiduvast vasest leidub tseruloplasmiini valguses. Kuigi sellel on üks sulfhüdrüülrühm, on plasmaalbumiini neutraalsetes lahustes vase esmane seondumiskoht valgumolekuli aminoots, mis sisaldab amiinlämmastikku, kahte deprotoneeritud peptiidlämmastikku ja ka küljes oleva imidasoolitsükli lämmastikku. kolmanda aminohappe ahel; kõik need lämmastikuaatomid kelaadivad vaske, moodustades tasapinnalise tsüklilise süsteemi. Hexaaqua-Cu 2+ muutub lämmastiku doonori aatomite arvu suurenedes tetragonaalsemaks (tasapinnalisemaks). Märkimisväärne kogus seedekulglasse sattunud vaske ärritab mao ja soolte närvilõpmeid ning põhjustab oksendamist. Ja vase krooniline liig põhjustab kasvu peatumist, hemolüüsi ja madalat hemoglobiinisisaldust, samuti koekahjustusi maksas, neerudes ja ajus. Enamikul patsientidel, kes põevad Wilsoni tõbe, mis on kaasasündinud ainevahetushäire, esineb tseruloplasmiini puudus. Sellistel patsientidel on maksas vasesisalduse tõus koos maksafunktsiooni häirega. Vase toksilisust saab vähendada MoO võtmisega.
Tsink. Inimestel on Zn 2+ ioon osa enam kui 20 metalliensüümist, sealhulgas ainevahetuses osalevatest nukleiinhapetest. Enamik veres leiduvaid Zn 2+ ioone leidub punastes verelibledes ensüümi karboanhüdraasi vajaliku kofaktorina. Tsingi puhul on teada ainult üks oksüdatsiooniaste lahuses. Zn 2+ roll ensüümi koostises on: a) kas substraadi otseses sidumises ja polarisatsioonis; b) kas kaudselt seotud vee või hüdroksiidioonide kaudu, nagu tavaliste happe-aluseliste katalüsaatorite ja nukleofiilide puhul. Suurem osa inimkehas leiduvast Zn 2+-st leidub lihastes ning suurim tsingi kontsentratsioon on sugunäärmes – eesnäärmes. Zn 2+ tase on homöostaasi kontrolli all. Tsingi puudust täheldatakse alkohoolikutel, aga ka arengumaade elanikel, kelle toidus on rohkesti kiudaineid ja viskoosseid toite. Tsingipuudus väljendub noortel nahahaigustes, kasvupeetuses, seksuaalarengu ja seksuaalfunktsioonide halvenemises. Kuigi inimestel pole afrodismi teada, on meeste normaalseks seksuaalkäitumiseks vajalik piisav kogus Zn 2+. Kuna inimese spermatogenees on mitmeetapiline protsess, nõuab häirete korrigeerimine ja seksuaaltervise taastamine Zn 2+ kontsentratsiooni suurendamise kaudu teatud aja. Tsingi lisamine võib häirida teiste metallide metaboolset tasakaalu, seetõttu tuleks selliseid sekkumisi läbi viia range meditsiinilise järelevalve all. Olgu seda nõuannet eriti rõhutatud, kuna hüpotees Zn 2+ /Cu 2+ suhtest kui peamisest südame isheemiatõve arengut põhjustavast tegurist (arteriaalse verevoolu lokaalne seiskumine) osutus üsna õigeks. Kahevalentse tsingi lisamine soodustab tsingipuudulikkusega patsientide haavade paranemist, kuid see ei aita, kui organismis on piisavas koguses Zn 2+. Lihas ja kalas on üsna palju tsinki, seega pole tööstusriikide elanikel toidulisandeid vaja; Lisaks võivad sellised lisandid olla ohtlikud, kui neid antakse kogustes, mis häirivad vase, raua ja muude oluliste metalliioonide imendumist.
Tsingisoolade liigne tarbimine võib põhjustada ägedaid soolehäireid, millega kaasneb iiveldus. Äge mürgistus selle elemendiga on tekkinud tsingitud (tsingitud) terasanumatesse pakendatud hapude puuviljamahlade tarbimisel. Kroonilise tsingimürgistuse juhtumid inimestel on üldiselt teadmata, kuid see võib tunduda hägune ja ebaselge. Näiteks kui tsink ja vask konkureerivad, võib tsingi liig põhjustada vase puudust, kui viimast esineb minimaalsetes kogustes. Samamoodi võib tsingi liig aeglustada loomade skeleti arengut, kui Ca ja P on minimaalses koguses. Üldiselt ei ole tsingiioon ohtlik ja ilmselt on selle mürgistuse peamine võimalus selle ühine esinemine mürgise kaadmiumiga (saastumise kujul).
Kaadmium. Üsna harva leidub kaadmiumi mineraalides ja pinnases koos tsingiga ligikaudu 0,1%. Sarnaselt tsingiga esineb see element ainult kahevalentse ioonina Cc1 2+. Kaadmiumiioon on suurem kui tsingiioon; see on oma suuruselt lähemal kaltsiumioonile, mis võimaldab seda kasutada nn Ca proovina. Kuid siiski on kaadmium ligandide sidumisvõime poolest rohkem sarnane tsingiga ja seetõttu täheldati tsingiga võrreldes mürgistuste arvu palju suuremates kogustes. Erinevalt Ca 2+ ioonist moodustavad nende metallide mõlemad ioonid tugevad sidemed ligandide doonorlämmastiku- ja väävliaatomitega. Liigne kaadmium häirib metallide ainevahetust, häirib tsingi ja teiste metallide ensüümide toimet, mis võib põhjustada tsingi ümberjaotumist organismis. Kaadmiumi toksilisuse täpne mehhanism pole teada, kuigi see on kindlasti mitmeastmeline.
Täielikult erinevalt CH 3 Hg + ioonist ei saa kaadmiumiioon kergesti platsentaarbarjääri ületada ja vastsündinutel puudub see element täielikult. Enamikul inimestel koguneb kaadmium toidust aeglaselt. Imendunud Cd 2+ vabaneb kehast väga aeglaselt, poolväärtusajaga üle 10 aasta. Selle tagajärjeks on kaadmiumisisalduse suurenemine neerudes kogu inimese eluea jooksul nullist sünnihetkel ligikaudu 20 mg-ni vanemas eas (mittesuitsetajatel) ja kuni 40 mg-ni täiskasvanud suitsetajatel. Suurem osa sellest elemendist on seotud metallotioneiiniga, mis on väikesed sulfhüdrüülasendajatega valgumolekulid, mille olemasolu ahelas stimuleerib kaadmium ise.
Äge kaadmiumimürgitus avaldub oksendamise, soolestiku spasmide ja peavaluna; see võib tekkida isegi joogiveest või muudest, eriti happelistest vedelikest, mis on Cd sisaldavate ühenditega kokku puutunud veetorudes, autodes või kaadmiumklaasidega nõudes. Kui kaadmium jõuab toiduga kehasse, transporditakse see verega teistesse organitesse, kus see seondub erütrotsüütide glutatiooni ja hemoglobiiniga. Suitsetajate veri sisaldab ligikaudu seitse korda rohkem kaadmiumi kui mittesuitsetajate veres. Krooniline kaadmiumimürgitus hävitab maksa ja neerud ning põhjustab raskeid neerufunktsiooni häireid. Kahjuks puudub kaadmiumimürgistuse raviks spetsiifiline ravi ja kelaativad ained suudavad kaadmiumi neerudesse ümber jaotada (mis on samuti ohtlik). Rikkalik tsingi, kaltsiumi, fosfaatide, D-vitamiini tarbimine ja valgudieet võivad kaadmiumimürgitust mõnevõrra vähendada. Kaadmiumimürgistuse eriti tõsist vormi on Jaapanis kirjeldatud kui "itai-itai" haigust (jaapani vaste "oh-oh"). Haiguse nimetus tuleneb osteomalaatsiaga kaasnevast valust seljas ja jalgades (tavaliselt eakatel naistel), mis põhjustab luude haprust (teatud juhtum, kus ühel inimesel on 72 luumurdu). Samuti täheldati tõsist neerufunktsiooni häiret proteinuuria (valgu ilmumine uriinis) tõttu, mis jätkus ka pärast kaadmiumiga kokkupuute lõpetamist. See haigus viib surma.
Elavhõbe on mürgine mis tahes kujul. Elavhõbeda ülemaailmne eraldumine gaaside kaudu maakoorest ja ookeanidest ületab vähemalt viis korda inimeste toodetud elavhõbeda kogust, kuid selle tööstuslik eraldumine on lokaalsem ja kontsentreeritum. Keskmine inimkeha sisaldab 13 mg elavhõbedat, mis ei anna mingit kasu. Varem kasutati raviainetena erinevaid elavhõbeda sooli (näiteks süüfilise ja gonorröa raviks kasutati elavhõbedabensoaati). Elavhõbeda reaktiivide kasutamine insektitsiidide ja fungitsiididena on põhjustanud kergeid ja raskeid mürgistusi, mis on tabanud tuhandeid inimesi. Seetõttu on elavhõbedamürgitus ülemaailmne probleem.
Elavhõbedat võib leida kolmel levinumal kujul ja ühel vähem levinud kujul, elavhõbeda ioonina Hg2+, mis jaguneb ebaproportsionaalselt elementaarseks elavhõbedaks ja kahevalentseks elavhõbedaks:
![]()
Selle reaktsiooni jaoks on tasakaalukonstandi väärtus
näitab, et reaktsioon kulgeb eelistatavalt paremalt vasakule. Kuid tegelikkuses kulgeb reaktsioon vasakult paremale tänu Hg 2+ iooni tugevale kompleksivõimele paljude ligandidega. Kolmas levinud elavhõbeda vorm on selle orgaaniline ühend metüülelavhõbe CH 3 Hg +.
Elavhõbe on toatemperatuuril vedel metall. Kuigi selle keemistemperatuur on 357 °C, on see väga lenduv ja seetõttu ohtlikum, kui üldiselt arvatakse. Üks kuupmeeter küllastunud (temperatuuril 25 °C) õhku sisaldab 20 mg Hg. See element on vees peaaegu lahustumatu; lahustuvuse piir 0,28 µM temperatuuril 25°C - 56 µg/l, s.o. 56 osa elavhõbedat miljardi osa vee kohta.
Mõlemad elavhõbeda katioonid (Hg 2+ ja metüülelavhõbe CH 3 Hg +) eelistavad lineaarset 2-koordinatsiooni. Nad moodustavad tugevamaid komplekse (kui enamik metalliioone) ligandidega, millel on üks doonor-aatom, eriti N või S. Ainult elavhõbe võib kõigist selles peatükis käsitletud metalliioonidest asendada vesinikku amiinides (kuid mitte ammooniumioonides) leeliselised lahused).
Tegelikult on sõna "merkaptaan" ise tuletatud elavhõbeda tugevast võimest seostuda tioolidega. Erütrotsüütides seostuvad Hg 2+ ioonid glutatiooni ja hemoglobiini sulfhüdrüülrühmadega segakompleksideks; Verre jääb ainult see elavhõbeda osa, mis tavaliselt inimkehas sisaldub. Hoolimata asjaolust, et Hg 2+ iooni toksilisuse molekulaarseks aluseks peetakse selle interaktsiooni sulfhüdrüülrühmadega, jääb teadmata, millised valgud läbivad metallatsiooni.
Hg 2+ ja CH 3 Hg + kiire vahetus doonorligandide, näiteks sulfhüdrüülrühmade üleliigse juuresolekul on toksikoloogias kriitilise tähtsusega. Just see määrab elavhõbeda kiire jaotumise kudedes sulfhüdrüülijääkide vahel. Veres jaotub CH 3 Hg' ioon samas proportsioonis nagu SH rühm: umbes 10% plasmas ja 90% erütrotsüütides, milles on nii hemoglobiini kui ka glutatiooni sulfhüdrüülrühmad. Elavhõbeda mõju tagasipööramiseks antakse elavhõbedamürgituse vastumürgina BAL-i (2,3-dimerkaptopropanooli), mis hõlbustab elavhõbeda ühtlast jaotumist kogu kehas; Kasutatakse ka hemodialüüsi kelaativate ainetega nagu tsüsteiin või L-atsetüülpenitsillamiin.
Sissehingamisel imendub elavhõbeda aur aktiivselt ja koguneb ajju, neerudesse ja munasarjadesse. Elavhõbe läbib platsentaarbarjääri; Äge mürgistus põhjustab kopsude hävimist. Keha kudedes muundatakse elementaarne elavhõbe iooniks, mis ühineb SH-rühmi sisaldavate molekulidega, sealhulgas valgu makromolekulidega. Krooniline elavhõbedamürgitus seisneb närvisüsteemi funktsioonide püsivas kahjustuses, põhjustab väsimust ja kõrgema mürgistuse korral iseloomulikku elavhõbedavärinat, kui väike värisemine katkeb märgatava värisemisega iga paari minuti tagant. Ainult 1 g elavhõbedasoola võtmine on surmav. Elavhõbeda soolad kogunevad neerudesse, kuid nad ei suuda sarnaselt elementaarsele elavhõbedale kiiresti vere- või platsentaarbarjääri läbida. Elavhõbeda allaneelamisel tekkiv äge mürgistus põhjustab valkude sadestumist seedetrakti limaskestadest, põhjustades valu, oksendamist ja kõhulahtisust. Kui patsient jääb ellu, on kriitiliseks organiks maks. Toimub teatav punaste vereliblede hemolüüs. Krooniline mürgistus väljendub kesknärvisüsteemi talitlushäiretes; Lewis Carrolli "Alice Imedemaal" tegelane Mad Hutter on ilmekas näide kutsehaiguse ohvrist karusnaha töötlemisel kasutatava Hg(N0 3) 2 soola mürgituse tõttu.
Orgaanilised elavhõbeda derivaadid, nagu metüülelavhõbedakloriid CH 3 HgCI, on oma lenduvuse tõttu väga mürgised. Elavhõbedat sisaldava saastunud vee mikroorganismid muudavad anorgaanilised elavhõbedaühendid kergesti monometüülelavhõbedaks CH 3 Hg +. Ja suurem osa kalade kehas olevast elavhõbedast on sellisel kujul, mis võib püsida aastaid. Kõrge CH 3 Hg + tase ei ole ilmselt nii mürgine kaladele kui inimestele, kelle aurude sissehingamisel või toidu allaneelamisel imenduvad CH 3 Hg + ioonid aktiivselt ja sisenevad punastesse verelibledesse, maksa ja neerudesse ning settivad aju (sealhulgas loote ajus), põhjustades kesknärvisüsteemi tõsist kumulatiivset pöördumatut düsfunktsiooni. Inimkehas on elavhõbeda poolväärtusaeg mitmest kuust mitme aastani. Toksiline toime võib olla varjatud ja mürgistusnähud võivad ilmneda alles mitu aastat hiljem.
Kaks kõige kuulsamat näidet massilisest elavhõbedamürgistusest olid põhjustatud CH 3 Hg + . 1956. aastal avastati Minamata tõbi Lõuna-Jaapanis, samanimelise merelahe lähedal. 1959. aastal näidati, et selle haiguse põhjuseks on elavhõbedaga mürgitatud kalade söömine kloriidina CH 3 HgCl, mille keemiatehas juhib otse lahe vetesse. Elavhõbeda kontsentratsioon oli nii kõrge, et kalad surid, kalu söönud linnud kukkusid otse merre ja mürgitatud toitu maitsenud kassid liikusid "tiirutades ja hüppades, siksakitades ja kokku kukkudes". Juba 1954. aastal vähendasid sellised “tantsud” siinset kassipopulatsiooni märgatavalt. Lahe vete elavhõbedareostuse mõõtmisi ei tehtud selles piirkonnas aga kuni 1959. aastani. Ja ainult tänu Jaapani iidsele kombele säilitada vastsündinute kuivanud nabanöör, sai võimalikuks tõestada, et lahe reostus elavhõbe sai alguse 1947. aastal. Kuid kuni 1968. aastani juhiti heitvett Laht ei peatatud!
Inimeste jaoks sai metüülelavhõbeda organismi sattumisest tingitud Minamata haigus alguse jäsemete ja näo tuimusest, naha tundlikkuse ja käte motoorse aktiivsuse halvenemisest näiteks kirjutamisel. Hiljem ilmnesid liigutuste koordinatsiooni puudumine, nõrkus, värisemine ja kõnnaku ebakindlus, samuti psüühikahäired, kõnehäired, kuulmine ja nägemine. Ja lõpuks üldine halvatus, jäsemete, eriti sõrmede deformatsioon, neelamisraskused, krambid ja surm. Traagiline on ka see, et selle haigusega vähe mõjutatud emade lapsed, kes ei pruukinud selle sümptomeid üldse tuvastada, surid tserebraalparalüüsi või muutusid idiootideks (tavaliselt ei seostata kesknärvihalvatus selgelt vaimse arengu mahajäämusega) . Ilmselt tungib ema kehas olev CH 3 Hg + läbi platsentaarbarjääri loote ülitundlikku kehasse. Naised haiguse tõsisemas staadiumis ei suutnud lapsi saada.
Tallium. Äärmiselt toksiliste talliumiühendite imendumine organismis põhjustab gastroenteriiti, perifeerset neuropaatiat ja sageli surma. Pikaajalise kroonilise talliumi kokkupuute korral täheldatakse kiilaspäisust. TI2SO4 kasutamine näriliste vastu on peatatud, kuna see on väga mürgine teistele kodu- ja metsloomadele. Talliumi peamine vorm kehas on T1 + ioon, kuigi T1C1 on kergelt lahustuv; Tallium esineb kehas ka T1 3+ kujul. Talliumiioonid ei ole palju suuremad kui kaalium, kuid need on palju mürgisemad ning talliumi läbilaskvus rakumembraanidest on sama, mis kaaliumil. Kuigi T1+ ja K+ ioonid on suuruselt sarnased, on esimene peaaegu neli korda paremini polariseeritav ja moodustab tugevaid komplekse. Näiteks toodab see lahustumatuid komplekse riboflaviiniga ja võib seetõttu häirida väävli metabolismi.
Pliid on tuntud peaaegu viis tuhat aastat ning Kreeka ja Araabia teadlased teadsid juba selle mürgisusest. Roomlastel oli kõrge pliimürgituse tase, kuna nad hoidsid veini ja küpsetasid toitu pliinõudes. Goya, nagu ka teised kunstnikud, kannatas sissehingamise ja juhusliku kokkupuute tõttu pliivärviga. Tänapäeval ohustab kõrge pliisisaldus linnalapsi, kuna nad puutuvad sageli kokku pliivärvidega värvitud esemetega, mängivad kasutatud patareidega ja meisterdavad ajakirjalehtedest (värvitrüki värvid sisaldavad 0,4% Pb-d) . Ja ennekõike põhjusel, et nad hingavad õhku, mis on saastunud autode heitgaasidest, mis sisaldavad tetraetüülplii Pb(C 2 H 5) 4 põlemisprodukte, mida lisatakse bensiinile, et tõsta kütuse oktaanarvu.
Peamine pliisaaste allikas on toit. Õnneks on allaneelatud plii imendumine madal, kuna tekivad lahustumatu fosfaat Pb 3 (P0 4) 2 ja aluseline karbonaat Pb 3 (C0 3) 2 (0H) 2. Imendunud plii koguneb luudesse, kus see seejärel osteoporoosi tõttu vabaneb, põhjustades hilinenud toksilisust. Tänapäeval sisaldab keskmine inimese geel umbes 120 mg pliid, s.o. kümneid kordi rohkem kui Egiptuse muumiatel. Sademeid põhjustavate ioonide puudumisel on pH = 7 juures plii Pb 2+ iooni kujul. Vastavalt rahvusvahelistele kokkulepetele ei tohiks plii sisaldus joogivees ületada 50 µg/l. Äge pliimürgitus põhjustab esmalt isukaotust ja oksendamist; Krooniline mürgistus põhjustab järk-järgult neerufunktsiooni häireid ja aneemiat.
Kontrollküsimused
- 1. Mis on metalliioonide bioanorgaanilise keemia uurimisobjekt ja teema?
- 2. Loetlege leelismetalliioonid (liitium, naatrium, kaalium, rubiidium, tseesium). Millised on nende ökoloogilised ja füsioloogilised põhiandmed?
- 3. Loetlege leelismuldmetallide (magneesium, kaltsium, baarium, strontsium, berüllium, lantaniidid) ioonid. Millised on nende ökoloogilised ja füsioloogilised põhiandmed?
- 4. Selgitage plii mõju inimorganismile. Milliseid meetmeid saab välja pakkuda inimeste tervise kaitsmiseks plii eest?
- 5. Kuidas kaadmium, elavhõbe ja arseen inimkehasse satuvad; milline on nende mõju?
- 6. Miks on seleeni tarbimine elusorganismile vajalik?
- 7. Määratleda bioanorgaaniline keemia ja näidata selle koht teiste keskkonnateaduste seas.
- 8. Defineerige mõisted "saasteaine" ja "ksenobiootiline aine". Nimetage tüüpilised raskmetallide rühma kuuluvad ksenobiootikumid.
- 9. 11miks soovitavad Moskva ja Moskva piirkonna arstid õpilastel ja koolilastel regulaarselt joodi sisaldavaid toite tarbida?
- 10. Nimetage raskmetallide aatomite peamised rändeteed atmosfääris ja hüdrosfääris.
- 11. Iseloomusta erinevaid rändevorme raskmetallide aatomite biosaadavuse seisukohalt.
- 12. Nimetage peamised keemilised protsessid, mis määravad raskemetallide aatomite esinemisvormid veekeskkonnas. Mis on mandrite pinnavete ja merevete raskemetallide aatomite geokeemia peamine erinevus?
- 13. Kuidas mõjutab huumusühendite esinemine vees raskmetallide aatomite biosaadavust? Nimeta biokeemilisi mehhanisme, mis kaitsevad elusorganisme (taimi ja loomi) raskmetallide aatomite toksilise toime eest.
- 14. Defineeri raskmetallid. Milline on nende roll biosfääris?
- 15. Kirjeldage kroomi ja elavhõbeda tsükleid.
- 16. Millised on keemiliste elementide jaotumise mustrid biosfääris?
- 17. Nimetage biosfääri tööstusliku saastamise keskkonnamõjud.
- 18. Määratlege suurimad lubatud kontsentratsioonid (kogused).
- 19. Kuidas teha kindlaks vee sobivus erinevateks otstarveteks?
- 20. Esitage toiduainetes sisalduvate saasteainete suurimate lubatud kontsentratsioonide väärtused.
Elumetallide mõiste. Naatrium ja kaalium. Aatomite struktuur ja katioonide hüdratatsiooni tunnused, mis määravad nende sisalduse rakuvälises ja rakusiseses keskkonnas.
Elu metallid– kümme elementi: K, Na, Ca, Mg, Mn, Fe, Co, Cu, Zn, Mo. Nende osakaal organismis moodustab 2,4%. Kõik kehas leiduvad elumetallid on kas vabade katioonide kujul või on kompleksi moodustavad ioonid, mis on seotud bioligandidega. Nad osalevad aktiivselt ainevahetuses.
Naatrium ja kaalium– groupIA elemendid. Selle rühma elementide aatomitel on väliskihis s-alamtasandil üks elektron, mille nad kipuvad andma ühendites partnerile, moodustades stabiilsed sümmeetrilised monokatioonid lähima väärisgaasi elektroonilise konfiguratsiooniga.
Elektroonilise struktuuri stabiilsuse ja madala positiivse laengutiheduse tõttu Na + ja K + katioonide pinnal ei saa nende välistasandi vabad aatomiasukad tõhusalt suhelda lähimate veemolekulide üksikute elektronpaaridega, mis Seetõttu säilivad need katiooni hüdratatsioonikihis ainult elektrostaatiliselt. Seetõttu ei hüdrolüüsi naatriumi- ja kaaliumikatioonid vesikeskkonnas ega kaldu praktiliselt moodustama komplekse.
Peamine erinevus naatriumi- ja kaaliumikatioonide omadustes on seotud nende pinnal oleva positiivse laengu tiheduse erinevusega: Na + katioonil on see kõrgem, mistõttu selle elektrostaatiline väli hoiab veemolekule tugevamana. Selle tulemusena on naatriumkatioonile iseloomulik positiivne hüdratatsioon ja kaaliumi katioonile negatiivne hüdratatsioon. See võib Valeri Ivanovitš Slesarevi sõnul selgitada, miks Na + ja K + katioonid on elussüsteemides antagonistid ja miks kaaliumi katioonid on valdavalt rakusisese ja naatriumkatioonid - rakkudevaheliste vedelike koostisosad.
K+ ioonide kontsentratsioon rakus on ligikaudu 35 korda suurem. Kui väljaspool seda, ja Na + ioonide kontsentratsioon rakuvälises vedelikus on 15 korda suurem kui raku sees. Paljude oluliste bioloogiliste protsesside läbiviimiseks on vaja pidevalt säilitada nende ioonide ebaühtlane jaotus, mis nõuab energiakulu, kuna ioonide ülekandmine läbi membraani peab toimuma nende kontsentratsiooni gradiendi vastu. See realiseerub kaaliumnaatriumpumba abil, mis ühe ATP molekuli hüdrolüüsi energia tõttu eemaldab rakust kolm Na + katiooni ja saadab rakku kaks K + katiooni. Ülekantud elektrilaengute tasakaalustamatuse tõttu on membraani sisepind laetud negatiivselt ja välispind positiivselt.
K-ioonide kõrge rakusisene kontsentratsioon tagab eelkõige rakusisese osmootse rõhu, ensümaatiliste süsteemide aktiveerimise ribosoomidel valkude sünteesiks ja süsivesikute oksüdatsiooniks. Erütrotsüütides osalevad K-ioonid hemoglobiini ja oksühemoglobiini puhversüsteemide töös ning aktiveerivad seeläbi ensüümi süsinikmonooksiidi karboanhüdraasi.
K+ ja Na+ ioonid aktiveerida rakumembraanide adenosiintrifosfataas (ATP-aas), mis annab energiat kaalium-naatriumpumbale. Need ioonid mõjutavad oluliselt kesknärvisüsteemi (KNS) aktiivsust. Na + ioonide liig ajukoore rakkudes põhjustab depressiooni, s.t. kesknärvisüsteemi aktiivsuse pärssimine. K-katioonide liig nendes rakkudes, vastupidi, erutab kesknärvisüsteemi, põhjustades maniakaalse seisundi.
Õpik: 338–341.
Magneesium ja kaltsium, aatomi struktuur ja nende ioonide hüdratatsiooni omadused. Magneesiumi ja kaltsiumi katioonide olemasolu vormid, asukoht ja roll organismis. Luukoe moodustumise ja hävimise reaktsioon ning selle funktsioon.
Täiskasvanu keha sisaldab umbes 20 g magneesiumi katioone ja 1000 g kaltsiumi. Pooled magneesiumi katioonidest ja peaaegu 99% kaltsiumist leidub luukoes, ülejäänud pehmetes kudedes. Magneesiumi katioonide päevane vajadus on umbes 0,3 g, kaltsiumi - 1 g ja naistel raseduse ajal suureneb kaltsiumi katioonide vajadus 3-4 korda.
Magneesium ja kaltsium on perioodilisuse tabeli IIA rühma elemendid. Selle rühma elementide aatomitel on nas-alataseme väliskihis kaks elektroni (12 Mg: 3s 2; 20 Ca: 4s 2), mis kipuvad olema antud ühendites partnerile.
Magneesiumi ja kaltsiumi katioonide omaduste erinevus vesikeskkonnas tuleneb nende pinnal oleva positiivse laengu tiheduse erinevusest. Kuna Mg 2+ katioonil on väiksem raadius kui Ca 2+ (vastavalt 66 ja 99), hüdraatub see paremini ning lisaks suudavad selle välise tasandi vabad aatomiorbitaalid, sealhulgas 3d– orbitaalid, suhelda üksikutega. veemolekulide elektronide paarid, moodustades üsna stabiilseid akvakomplekse 2+.
Magneesiumikatioon on kaltsiumkatiooniga võrreldes võimeline moodustama kovalentseid sidemeid. Sellega seoses on magneesiumikatioonid erinevalt kaltsiumi katioonidest võimelised hüdrolüüsima:
Mg2+ +H2O⇌ Mg(OH)+ + H+
Suur osa väljaspool luid paiknevatest magneesiumi katioonidest on koondunud rakkudesse. Magneesiumioonid mängivad olulist rolli osmootse rõhu säilitamisel rakkude sees. Magneesiumi põhiosa veres sisaldub ioniseeritud kujul, s.o. vee kujul (55–60%) on ligikaudu 30% seotud valkudega ja 10–15% fosfolipiidide ja nukleotiididega kompleksühenditest.
Komplekside moodustumise tõttu on magneesiumi katioonid üks peamisi ensümaatiliste protsesside aktivaatoreid. Seega aktiveerivad nad oksüdatiivse fosforüülimise, DNA replikatsiooni ja luu mineralisatsiooni ensüüme.
Erinevalt magneesiumioonidest on kaltsiumi katioonid koondunud valdavalt rakkudevahelistesse vedelikesse. Kaltsiumi metabolismi kontrollivad kõrvalkilpnäärme ja kilpnäärme hormoonid, samuti D-vitamiin.
Luukoe peamine mineraalne komponent on kaltsiumvesinikfosfaat
Ca 5 (PO 4) 3 OH (hüdroksüapatiit). Luukoe tagab Ca 2+ ioonide kontsentratsiooni hoidmise bioloogilistes vedelikes teatud tasemel, seega võib seda pidada organismi kaltsiumipuhvriks.
Kompaktne luukude (kompaktne aine) on üks kahest luukoe tüübist, mis moodustavad luu. Tagab luu toetavad, kaitsvad funktsioonid, toimib keemiliste elementide hoidlana.
Kompaktne aine moodustab enamiku luude ajukoore. See on palju tihedam, raskem ja tugevam kui käsnjas aine. Kompaktne luukude moodustab umbes 80% inimese luustiku kogumassist. Kompaktse aine peamine struktuurne ja funktsionaalne üksus on osteoon.
Õpik: 341 – 344.
Raud ja koobalt, aatomi struktuur ja iseloomulikud oksüdatsiooniastmed. Nende metallide ühendite happe-aluse, redoks- ja kompleksi moodustavad omadused. Nende metallide ühendite roll elusorganismis.
Inimkeha sisaldab umbes 5 g rauda ja 1,2 mg koobaltit. Suurem osa rauast (70%) on kontsentreeritud vere hemoglobiinis; 14% koobaltist leidub luudes, 43% lihastes ja ülejäänud pehmetes kudedes. Päevane raua tarbimine on 10-20 mg ja koobalti 0,3 mg.
Raud ja koobalt– elektrooniliste konfiguratsioonidega perioodilisustabeli 4. perioodi VIIIB rühma elemendid 26 Fe: 1s 2 2s 2 2p 6 3s 2 3p 6 4s 2 ; 27 Co: 1 s 2 2 s 2 2p 6 3s 2 3p 6 3p 7 4s 2
Raua ja koobalti kõige iseloomulikumad oksüdatsiooniastmed +2 ja +3.
Vesilahustes on Fe 2+, Fe 3+, Co 2+ ja Co 3+ katioonid hüdraatunud, moodustades kuue koordinatsiooniga veekompleksid.
Fe 2+ on tugev redutseerija, mida võib oksüdeerida isegi õhuhapnik.
Co 3+ on nii tugev oksüdeerija, et oksüdeerib isegi vett:
4Fe(OH)2 + O2 + 2H2O = 4Fe(OH)3
2Co 2 (SO 4) 3 + 2H 2 O = 4 CoSO 4 + 2H 2 SO 4 + O 2
Raua ja koobalti oksiididel ja hüdroksiididel, olenemata oksüdatsiooniastmest, on nõrgad amfoteersed omadused, kusjuures põhiomadused on ülekaalus, eriti kahevalentse oleku korral, kui interaktsioon toimub ainult leeliste kontsentreeritud lahustega ja kuumutamisel.
Raua ja koobalti katioonid on väga altid komplekside moodustumisele. Tõenäoliselt nende jaoks kooskõlastusnumber kuus: 

Kompleks raua ja koobalti katioonid tugevalt, kuid mõjutavad erinevalt nende redoksomadusi sõltuvalt oksüdeeritud ja redutseeritud vormide komplekside stabiilsuse suhtest samade ligandidega.
Kompleks Co 3+, mille ligandid on veemolekulidest aktiivsemad, muudab selle vesilahustes stabiilseks.
Koobalt, üks organismile elutähtsatest mikroelementidest. See on osa vitamiinist B12 (kobalamiin). Koobalt osaleb hematopoeesis, närvisüsteemi ja maksa funktsioonides ning ensümaatilistes reaktsioonides. Inimkeha sisaldab 0,2 mg koobaltit inimese iga kilogrammi kohta. Koobalti puudumisel areneb akobaltoos.
Elusorganismides raud on oluline mikroelement, mis katalüüsib hapnikuvahetuse (hingamise) protsesse. Tavaliselt siseneb raud ensüümidesse kompleksi kujul, mida nimetatakse heemiks. Eelkõige on see kompleks hemoglobiinis, mis on kõige olulisem valk, mis tagab hapniku transpordi veres kõigisse inimeste ja loomade organitesse. Ja just tema värvib vere iseloomulikku punast värvi.
Seal on suur seltskond umbes 50 liiki, rauda sisaldavad ensüümid - tsütokroomid, mis katalüüsivad elektronide ülekande protsessi hingamisahelas, muutes raua oksüdatsiooniastet Fe 3+ + e - Fe 2+
Õpik: 349 – 352.
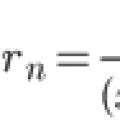 Taylori seeria laiendus Cauchy probleemi ligikaudne lahendus tavalisele
Taylori seeria laiendus Cauchy probleemi ligikaudne lahendus tavalisele Mida koolis EI õpetata, mida koolis ei õpetata, küsi
Mida koolis EI õpetata, mida koolis ei õpetata, küsi Raha mõtlemise valem (A
Raha mõtlemise valem (A