The problem of localization of functions in the cerebral cortex. Localization of functions in the cerebral cortex
The cerebral cortex is the evolutionarily youngest formation, which in humans has reached its greatest values in relation to the rest of the brain mass. In humans, the mass of the cerebral cortex is on average 78% of the total mass of the brain. The cerebral cortex is extremely important in the regulation of the body’s vital functions, the implementation of complex forms of behavior and the development of neuropsychic functions. These functions are provided not only by the entire mass of the cortical substance, but also by the unlimited possibilities of associative connections between the cells of the cortex and subcortical formations, which creates conditions for the most complex analysis and synthesis of incoming information, for the development of forms of learning that are inaccessible to animals.
Speaking about the leading role of the cerebral cortex in neurophysiological processes, we should not forget that this higher department can function normally only in close interaction with subcortical formations. The contrast between the cortex and underlying parts of the brain is largely schematic and conditional. IN last years ideas about the vertical organization of the functions of the nervous system and about circular cortical-subcortical connections are developing.
The cells of the cortex are specialized to a much lesser extent than the nuclei of the subcortical formations. It follows that the compensatory capabilities of the cortex are very high - the functions of the affected cells can be taken over by other neurons; damage to fairly large areas of the cortex can clinically appear very blurred (the so-called clinical silent zones). The absence of narrow specialization of cortical neurons creates conditions for the emergence of a wide variety of interneuron connections, the formation of complex “ensembles” of neurons that regulate various functions. This is the most important basis of learning ability. The theoretically possible number of connections between the 14 billion cells of the cerebral cortex is so large that during a person’s life a significant part of them remains unused. This once again confirms the unlimited possibilities of human learning.
Despite the known nonspecificity of cortical cells, certain groups of them are anatomically and functionally more closely related to certain specialized parts of the nervous system. The morphological and functional ambiguity of different areas of the cortex allows us to speak about the cortical centers of vision, hearing, touch, etc., which have a specific localization. In the works of researchers of the 19th century, this principle of localization was taken to the extreme: attempts were made to identify centers of will, thinking, the ability to understand art, etc. At present, it would be incorrect to talk about the cortical center as a strictly limited group of cells. It should be noted that the specialization of nerve links is formed in the process of life.
According to I.P. Pavlov, the brain center, or the cortical part of the analyzer, consists of a “core” and “scattered elements.” The “nucleus” is a relatively morphologically homogeneous group of cells with a precise projection of receptor fields. “Scattered elements” are located in a circle or at a certain distance from the “core”: they carry out a more elementary and less differentiated analysis and synthesis of incoming information.
Of the 6 layers of cortical cells, the upper layers are most developed in humans compared to similar layers in animals and are formed in ontogenesis much later than the lower layers. The lower layers of the cortex have connections with peripheral receptors (layer IV) and with muscles (layer V) and are called “primary” or “projection” cortical zones due to their direct connection with the peripheral parts of the analyzer. Above the “primary” zones are built systems of “secondary” zones (layers II and III), in which associative connections with other parts of the cortex predominate, therefore they are also called projection-associative.
Thus, two groups of cellular zones are identified in the cortical representations of the analyzers. Such a structure is found in the occipital zone, where the visual pathways are projected, in the temporal zone, where the auditory pathways end, in the posterior central gyrus - the cortical section of the sensitive analyzer, in the anterior central gyrus - the cortical motor center. The anatomical heterogeneity of the “primary” and “secondary” zones is accompanied by physiological differences. Experiments with stimulation of the cortex have shown that stimulation of the primary zones of the sensory regions leads to the emergence of elementary sensations. For example, irritation of the occipital regions causes a sensation of flickering light points, lines, etc. With irritation of the secondary zones, more complex phenomena arise: the subject sees variously designed objects - people, birds, etc. It can be assumed that it is in the secondary zones that operations are carried out gnosis and partly praxis.
In addition, tertiary zones, or zones of overlap of cortical representations of individual analyzers, are distinguished in the cortex. In humans, they occupy a very significant place and are located primarily in the parieto-temporo-occipital region and in the frontal zone. Tertiary zones enter into extensive connections with cortical analyzers and thereby ensure the development of complex, integrative reactions, among which meaningful actions occupy the first place in humans. In the tertiary zones, therefore, planning and control operations take place, requiring the complex participation of different parts of the brain.
In early childhood, the functional zones of the cortex overlap each other, their boundaries are diffuse, and only in the process practical activities There is a constant concentration of functional zones into delineated centers separated from each other. In the clinic, adult patients experience very constant symptom complexes when certain areas of the cortex and associated nerve pathways are affected.
In childhood, due to incomplete differentiation of functional zones, focal damage to the cerebral cortex may not have a clear clinical manifestation, which should be remembered when assessing the severity and boundaries of brain damage in children.
In functional terms, we can distinguish the main integrative levels of cortical activity.
The first signaling system is associated with the activities of individual analyzers and carries out the primary stages of gnosis and praxis, i.e., the integration of signals arriving through the channels of individual analyzers, and the formation of response actions taking into account the state of external and internal environment, as well as past experience. This first level includes visual perception of objects with concentration of attention on certain of its details, voluntary movements with active strengthening or inhibition of them.
A more complex functional level of cortical activity unites the systems of various analyzers, includes a second signal system), unites the systems of various analyzers, making it possible for a meaningful perception of the environment, an attitude towards the surrounding world “with knowledge and understanding.” This level of integration is closely related to speech activity, and the understanding of speech (speech gnosis) and the use of speech as a means of address and thinking (speech praxis) are not only interconnected, but also determined by various neurophysiological mechanisms, which is of great clinical importance.
The highest level of integration is formed in a person in the process of his maturation as a social being, in the process of mastering the skills and knowledge that society has.
The third stage of cortical activity plays the role of a kind of dispatcher of complex processes of higher nervous activity. It ensures the purposefulness of certain acts, creating conditions for their best implementation. This is achieved by “filtering” signals that are currently of greatest importance from signals of secondary importance, performing probabilistic forecasting of the future and forming long-term tasks.
Of course, complex cortical activity could not be carried out without the participation of the information storage system. Therefore, memory mechanisms are one of the most important components of this activity. In these mechanisms, not only the functions of recording information (memorization), but also the functions of obtaining the necessary information from memory “storages” (memory), as well as the functions of transferring information flows from blocks of RAM (what is needed at the moment) into long-term memory blocks and vice versa. Otherwise, it would be impossible to learn new things, since old skills and knowledge would interfere with this.
Recent neurophysiological studies have made it possible to establish which functions are predominantly characteristic of certain parts of the cerebral cortex. Even in the last century, it was known that the occipital region of the cortex is closely connected with the visual analyzer, the temporal region - with the auditory (Heschl's gyrus), taste analyzer, the anterior central gyrus - with the motor, and the posterior central gyrus - with the musculocutaneous analyzer. We can conditionally assume that these departments are associated with the first type of cortical activity and provide the simplest forms of gnosis and praxis.
Parts of the cortex located in the parietotemporal-occipital region take an active part in the formation of more complex gnostic-praxic functions. Damage to these areas leads to more complex forms of disorders. Wernicke's Gnostic speech center is located in the temporal lobe of the left hemisphere. The motor speech center is located somewhat anterior to the lower third of the anterior central gyrus (Broca's center). In addition to the centers of oral speech, there are sensory and motor centers of written speech and a number of other formations, one way or another related to speech. The parieto-temporo-occipital region, where the pathways coming from various analyzers close, is of utmost importance for the formation of higher mental functions. The famous neurophysiologist and neurosurgeon W. Penfield called this area the interpretive cortex. In this area there are also formations involved in memory mechanisms.
Particular importance is attached to the frontal region. According to modern concepts, it is this section of the cerebral cortex that takes an active part in organizing purposeful activity, in long-term planning and determination, i.e. it belongs to the third type of cortical functions.
The main centers of the cerebral cortex. Frontal lobe. The motor analyzer is located in the anterior central gyrus and paracentral lobule (Brodmann's areas 4, 6 and 6a). In the middle layers there is an analyzer of kinesthetic stimuli coming from skeletal muscles, tendons, joints and bones. In layer V and partly VI, giant pyramidal cells of Betz are located, the fibers of which form the pyramidal path. The anterior central gyrus has a certain somatotopic projection and is connected with the opposite half of the body. The muscles of the lower extremities are projected in the upper parts of the gyrus, and the muscles of the face in the lower parts. The trunk, larynx, and pharynx are represented in both hemispheres (Fig. 55).
The center of rotation of the eyes and head in the opposite direction is located in the middle frontal gyrus in the premotor area (fields 8, 9). The work of this center is closely connected with the system of the posterior longitudinal fasciculus, the vestibular nuclei, formations of the striopallidal system, which is involved in the regulation of torsion, as well as with the cortical part of the visual analyzer (field 17).
In the posterior parts of the superior frontal gyrus there is a center that gives rise to the fronto-pontocerebellar pathway (field 8). This area of the cerebral cortex is involved in ensuring the coordination of movements associated with upright posture, maintaining balance while standing and sitting, and regulates the work of the opposite hemisphere of the cerebellum.
The motor speech center (speech praxis center) is located in the posterior part of the inferior frontal gyrus - Broca's gyrus (area 44). The center provides analysis of kinesthetic impulses from the muscles of the speech-motor apparatus, storage and implementation of “images” of speech automatisms, the formation of oral speech, and is closely connected with the location posterior to it of the lower part of the anterior central gyrus (projection zone of the lips, tongue and larynx) and with that located in front of it musical motor center.
The musical motor center (field 45) provides a certain tonality, modulation of speech, as well as the ability to compose musical phrases and sing.
The center of written speech is localized in the posterior part of the middle frontal gyrus in close proximity to the projection cortical area of the hand (field 6). The center ensures the automaticity of writing and is functionally connected with Broca's center.
Parietal lobe. The center of the skin analyzer is located in the posterior central gyrus of fields 1, 2, 3 and the cortex of the superior parietal region (fields 5 and 7). In the posterior central gyrus, tactile, pain, and temperature sensitivity of the opposite half of the body is projected. The sensitivity of the leg is projected in the upper sections, and the sensitivity of the face is projected in the lower sections. Boxes 5 and 7 represent elements of deep sensitivity. Posterior to the middle sections of the posterior central gyrus is the center of stereognosis (fields 7,40 and partly 39), which provides the ability to recognize objects by touch.
Posterior to the upper parts of the posterior central gyrus is a center that provides the ability to recognize one’s own body, its parts, their proportions and relative positions (field 7).
The center of praxis is localized in the inferior parietal lobule on the left, the supramarginal gyrus (fields 40 and 39). The center provides storage and implementation of images of motor automatisms (praxis functions).
In the lower parts of the anterior and posterior central gyri there is the center of the analyzer of interoceptive impulses of internal organs and blood vessels. The center has close connections with subcortical vegetative formations.
Temporal lobe. The center of the auditory analyzer is located in the middle part of the superior temporal gyrus, on the surface facing the insula (Heschl's gyrus, areas 41, 42, 52). These formations provide the projection of the cochlea, as well as the storage and recognition of auditory images.
The center of the vestibular analyzer (fields 20 and 21) is located in the lower parts of the outer surface of the temporal lobe, is projection, and is in close connection with the lower basal parts of the temporal lobes, giving rise to the occipitotemporal cortical-pontine-cerebellar pathway.
Rice. 55. Scheme of localization of functions in the cerebral cortex (A - D). I - projection motor zone; II - center of rotation of the eyes and head in the opposite direction; III - projection sensitivity zone; IV - projection visual zone; projection gnostic zones: V - hearing; VI - smell, VII - taste, VIII - gnostic zone of the body diagram; IX - stereognosis zone; X - gnostic visual zone; XI - Gnostic reading zone; XII - gnostic speech zone; XIII - praxis zone; XIV - praxic speech zone; XV - practical writing zone; XVI - zone of control over the function of the cerebellum.
The center of the olfactory analyzer is located in the phylogenetically most ancient part of the cerebral cortex - in the hook and ammon's horn (field 11a, e) and provides projection function, as well as storage and recognition of olfactory images.
The center of the taste analyzer is located in the immediate vicinity of the center of the olfactory analyzer, i.e. in the hook and ammon's horn, but, in addition, in the lowest part of the posterior central gyrus (area 43), as well as in the insula. Like the olfactory analyzer, the center provides projection function, storage and recognition of taste images.
The acoustic-gnostic sensory speech center (Wernicke's center) is localized in the posterior parts of the superior temporal gyrus on the left, in the depth of the lateral sulcus (area 42, as well as areas 22 and 37). The center provides recognition and storage of sound images of oral speech, both one’s own and others’.
In the immediate vicinity of Wernicke's center (the middle third of the superior temporal gyrus - area 22) there is a center that ensures the recognition of musical sounds and melodies.
Occipital lobe. The center of the visual analyzer is located in the occipital lobe (fields 17, 18, 19). Field 17 is a projection visual zone, fields 18 and 19 provide storage and recognition of visual images, visual orientation in an unusual environment.
On the border of the temporal, occipital and parietal lobes is the center of the written speech analyzer (field 39), which is closely connected with the Wernicke center of the temporal lobe, with the center of the visual analyzer of the occipital lobe, as well as with the centers of the parietal lobe. The reading center provides recognition and storage of written language images.
Data on the localization of functions were obtained either as a result of irritation of various parts of the cortex in an experiment, or as a result of the analysis of disturbances arising as a result of damage to certain areas of the cortex. Both of these approaches can only indicate the participation of certain cortical zones in certain mechanisms, but do not at all mean their strict specialization or unambiguous connection with strictly defined functions.
In the neurological clinic, in addition to signs of damage to areas of the cerebral cortex, there are symptoms of irritation of its individual areas. In addition, in childhood, phenomena of delayed or impaired development of cortical functions are observed, which significantly modifies the “classical” symptoms. The existence of different functional types of cortical activity causes different symptoms of cortical lesions. Analysis of these symptoms allows us to identify the nature of the lesion and its location.
Depending on the types of cortical activity, it is possible to distinguish among cortical lesions disturbances of gnosis and praxis on different levels integration; speech disorders due to their practical importance; disorders of regulation of purposefulness, purposefulness of neurophysiological functions. With each type of disorder, the memory mechanisms involved in a given functional system may also be disrupted. In addition, more total memory impairment is possible. In addition to relatively local cortical symptoms, more diffuse symptoms are also observed in the clinic, manifesting primarily in intellectual disability and behavioral disorders. Both of these disorders are of particular importance in child psychiatry, although in essence many variants of such disorders can be considered borderline between neurology, psychiatry and pediatrics.
The study of cortical functions in childhood has a number of differences from the study of other parts of the nervous system. It is important to establish contact with the child and maintain a relaxed tone of conversation with him. Since many diagnostic tasks presented to a child are very complex, one must strive to ensure that he not only understands the task, but also becomes interested in it. Sometimes when examining children who are overly distracted, motorically disinhibited, or mentally retarded, a lot of patience and ingenuity must be applied to identify existing abnormalities. In many cases, the analysis of a child’s cortical functions is helped by parents’ reports about his behavior at home, at school, and school characteristics.
When studying cortical functions, it is important psychological experiment, the essence of which is the presentation of standardized, targeted tasks. Certain psychological methods allow one to evaluate certain aspects of mental activity in isolation, while others allow them to be assessed more comprehensively. These include so-called personality tests.
Gnosis and its disorders. Gnosis literally means recognition. Our orientation in the surrounding world is associated with recognizing the shape, size, spatial relationship of objects and, finally, understanding their meaning, which is contained in the name of the object. This stock of information about the surrounding world consists of the analysis and synthesis of sensory impulse flows and is stored in memory systems. The receptor apparatus and the transmission of sensory impulses with lesions of higher gnostic mechanisms are preserved, but the interpretation of these impulses and the comparison of the received data with images stored in memory are disrupted. As a result, a disorder of gnosis occurs - agnosia, the essence of which is that while the perception of objects is preserved, the feeling of their “familiarity” is lost and the world, previously so familiar in detail, becomes alien, incomprehensible, devoid of meaning.
But gnosis cannot be imagined as a simple comparison, recognition of an image. Gnosis is a process of continuous updating, clarification, concretization of the image stored in the memory matrix, under the influence of its repeated comparison with the received information.
Total agnosia, in which complete disorientation is observed, is rare. Much more often, gnosis is disrupted in any one analytical system, and depending on the degree of damage, the severity of agnosia varies.
Visual agnosia occur when the occipital cortex is damaged. The patient sees the object, but does not recognize it. There may be various options here. In some cases, the patient correctly describes the external properties of an object (color, shape, size), but cannot recognize the object. For example, a patient describes an apple as “something round and pink,” without recognizing the apple as an apple. But if you give this object to the patient, he will recognize it when he feels it. There are times when the patient does not recognize familiar faces. Some patients with a similar disorder are forced to remember people based on some other characteristics (clothing, mole, etc.). In other cases of agnosia, the patient recognizes an object, names its properties and function, but cannot remember what it is called. These cases belong to the group of speech disorders.
In some forms of visual agnosia, spatial orientation and visual memory are impaired. In practice, even if an object is not recognized, we can talk about violations of memory mechanisms, since the perceived object cannot be compared with its image in the Gnostic matrix. But there are also cases when, when an object is presented again, the patient says that he has already seen it, although he still cannot recognize it. If spatial orientation is impaired, the patient not only does not recognize previously familiar faces, houses, etc., but can also walk in the same place many times without knowing it.
Often, with visual agnosia, recognition of letters and numbers also suffers, and loss of reading ability occurs. The isolated type of this disorder will be analyzed in the analysis of speech function.
To study visual gnosis, a set of objects is used. Presenting them to the subject, they are asked to identify and describe them. appearance, compare which objects are larger and which are smaller. They also use a set of pictures, color, plain and outline. They evaluate not only the recognition of objects, faces, but also plots. At the same time, you can test visual memory: present several pictures, then mix them with previously unseen ones and ask the child to choose familiar pictures. At the same time, work time, persistence, and fatigue are also taken into account.
It should be borne in mind that children recognize contour pictures worse than colored and monochromatic ones. Understanding the plot is related to the child’s age and degree of mental development. At the same time, agnosia in the classical form is rare in children due to incomplete differentiation of cortical centers.
Auditory agnosia. They occur when the temporal lobe is damaged in the area of Heschl’s gyrus. The patient cannot recognize previously familiar sounds: the ticking of a clock, the ringing of a bell, the sound of flowing water. Possible impairment of recognition of musical melodies - amusia. In some cases, the determination of the direction of sound is disrupted. In some types of auditory agnosia, the patient is unable to distinguish the frequency of sounds, such as metronome beats.
Sensitive agnosia are caused by impaired recognition of tactile, pain, temperature, proprioceptive images or their combinations. They occur when the parietal region is damaged. This includes astereognosis, body diagram disorders. In some variants of astereognosis, the patient not only cannot identify an object by touch, but is also unable to determine the shape of the object or the features of its surface. Sensitive agnosia also includes anosognosia, in which the patient is not aware of his defect, for example, paralysis. Phantom sensations can be attributed to disorders of sensitive gnosis.
When examining children, it should be borne in mind that a small child cannot always show parts of his body correctly; The same applies to patients suffering from dementia. In such cases, there is, of course, no need to talk about a disorder of the body diagram.
Taste and olfactory agnosia are rare. In addition, recognition of odors is very individual and is largely related to personal experience person.
Praxis and its disorders. Praxis refers to purposeful action. A person learns a lot of special motor acts in the course of life. Many of these skills, being formed with the participation of higher cortical mechanisms, are automated and become the same integral human ability as simple movements. But when the cortical mechanisms involved in the implementation of these acts are damaged, peculiar movement disorders arise - apraxia, in which there is no paralysis, no disturbances of tone or coordination, and even simple voluntary movements are possible, but more complex, purely human motor acts are disrupted. The patient suddenly finds himself unable to perform such seemingly simple actions as shaking hands, fastening buttons, combing his hair, lighting a match, etc. Apraxia occurs primarily when the parieto-temporo-occipital region of the dominant hemisphere is affected. In this case, both halves of the body are affected. Apraxia can also occur with damage to the subdominant right hemisphere (in right-handed people) and the corpus callosum, which connects both hemispheres. In this case, apraxia is detected only on the left. With apraxia, the plan of action suffers, i.e., the formation of a continuous chain of motor automatisms. Here it is appropriate to quote the words of K. Marx: “Human action differs from the work of the “best bee” in that before building, a person has already built in his head. At the end of the labor process, a result is obtained that was already ideal before the start of this process, that is, in the mind of the worker.”
Due to a violation of the action plan, when trying to complete a task, the patient makes many unnecessary movements. In some cases, parapraxia is observed when an action is performed that is only vaguely reminiscent of the given task. Sometimes perseverations are also observed, i.e. getting stuck on some actions. For example, the patient is asked to make an inviting movement with his hand. After completing this task, they offer to wag their finger, but the patient still performs the first action.
In some cases, with apraxia, ordinary, everyday actions are preserved, but professional skills are lost (for example, the ability to use a plane, screwdriver, etc.).
According to clinical manifestations, several types of apraxia are distinguished: motor, ideational and constructive.
Motor apraxia. The patient cannot perform actions according to instructions or even imitation. He is asked to cut paper with scissors, lace a shoe, line paper with a pencil and ruler, etc., but the patient, although he understands the task, cannot complete it, showing complete helplessness. Even if you show how this is done, the patient still cannot repeat the movement. In some cases, it turns out to be impossible to perform such simple actions as squatting, turning, clapping your hands.
Ideatorial apraxia. The patient cannot perform actions on a task with real and imaginary objects (for example, show how to comb one's hair, stir sugar in a glass, etc.), while at the same time the actions of imitation are preserved. In some cases, the patient can automatically perform certain actions without thinking. For example, he purposefully cannot fasten a button, but performs this action automatically.
Constructive apraxia. The patient can perform various actions by imitation and by verbal orders, but is unable to create a qualitatively new motor act, put together a whole from parts, for example, make a certain figure out of matches, put together a pyramid, etc.
Some variants of apraxia are associated with impaired gnosis. The patient does not recognize the object or his body diagram is disturbed, so he is unable to perform tasks or performs them uncertainly and not entirely correctly.
To study praxis, a number of tasks are offered (sit down, shake a finger, comb your hair, etc.). They are also presented with tasks for actions with imaginary objects (they are asked to show how they eat, how they make phone calls, how they cut wood, etc.). Assess how the patient can imitate the actions shown.
Special psychological techniques are also used to study gnosis and praxis. Among them, an important place is occupied by Seguin boards with recesses of various shapes, into which you need to insert figures corresponding to the recesses. This method also allows you to assess the degree of mental development. The Koss technique is also used: a set of cubes of different colors. From these cubes you need to put together a pattern that matches the one shown in the picture. Older children are also offered a Link cube: they need to fold a cube out of 27 differently colored cubes so that all its sides are the same color. The patient is shown the assembled cube, then they destroy it and ask him to put it back together.
In these techniques great importance has how the child performs the task: whether he acts by trial and error or according to a specific plan.
Rice. 56. Diagram of connections of speech centers and regulation of speech activity.
1 - writing center; 2 - Broca's center; 3 - center of praxis; 4 - center of proprioceptive gnosis; 5 - reading center; 6 - Wernicke center; 7 - center of auditory gnosis; 8 - center of visual gnosis.
It is important to remember that praxis develops as the child matures, so young children cannot yet perform such simple actions as combing their hair, fastening buttons, etc. Apraxia in its classic form, like agnosia, occurs mainly in adults.
Speech and its disorders. IN Visual, auditory, motor and kinesthetic analyzers take part in the implementation of speech functions, as well as writing and reading. Of great importance are the preservation of the innervation of the muscles of the tongue, larynx, soft palate, the condition of the paranasal sinuses and the oral cavity, which play the role of resonator cavities. In addition, coordination of breathing and pronunciation of sounds is important.
For normal speech activity, the coordinated functioning of the entire brain and other parts of the nervous system is necessary. Speech mechanisms have a complex and multi-stage organization (Fig. 56).
Speech is the most important human function, therefore, cortical speech zones located in the dominant hemisphere (Broca's and Wernicke's centers), motor, kinetic, auditory and visual areas, as well as afferent and efferent pathways related to the pyramidal and extrapyramidal systems take part in its implementation. , analyzers of sensitivity, hearing, vision, bulbar parts of the brain, visual, oculomotor, facial, auditory, glossopharyngeal, vagus and hypoglossal nerves.
The complexity and multi-stage nature of speech mechanisms also determines the variety of speech disorders. When the innervation of the speech apparatus is disrupted, dysarthria- articulation disorder, which may be caused by central or peripheral paralysis of the speech-motor apparatus, damage to the cerebellum, or striopallidal system.
There are also dyslalia- phonetically incorrect pronunciation individual sounds. Dyslalia can be functional in nature and can be quite successfully eliminated with speech therapy sessions. Under alalia understand the delay speech development. Usually to V.A. At the age of 10 years, the child begins to speak, but sometimes this happens much later, although the child understands speech addressed to him well. Delayed speech development also affects mental development, since speech is the most important means of information for a child. However, there are also cases of alalia associated with dementia. The child is lagging behind in mental development, and therefore his speech is not formed. These different cases of alalia need to be differentiated, as they have different prognoses.
With the development of speech function in the dominant hemisphere (in the left for right-handers, in the right for left-handers), gnostic and practical speech centers are formed, and subsequently - writing and reading centers.
Cortical speech disorders are variants of agnosia and apraxia. There are expressive (motor) and impressive (sensory) speech. Cortical motor speech disorder is apraxia of speech, sensory speech - speech agnosia. In some cases, the recall of the necessary words is impaired, i.e., memory mechanisms suffer. Speech agnosia and apraxia are called aphasia.
It should be remembered that speech disorders can be a consequence of general apraxia (apraxia of the trunk, limbs) or oral apraxia, in which the patient loses the ability to open his mouth, puff out his cheeks, and stick out his tongue. These cases are not aphasias; speech apraxia here arises secondarily as a manifestation of general praxic disorders.
Speech disorders in childhood, depending on the causes of their occurrence, can be divided into the following groups:
I. Speech disorders associated with organic damage to the central nervous system. Depending on the level of damage to the speech system, they are divided into:
1) aphasia—decay of all components of speech as a result of damage to cortical speech areas;
2) alalia - systemic underdevelopment of speech due to lesions of cortical speech zones in the pre-speech period;
3) dysarthria - a violation of the sound-pronunciation aspect of speech as a result of a violation of the innervation of the speech muscles.
Depending on the location of the lesion, several forms of dysarthria are distinguished.
II. Speech disorders associated with functional changes
central nervous system:
1) stuttering;
2) mutism and surdomutism.
III. Speech disorders associated with defects in the structure of the articulatory apparatus (mechanical dyslalia, rhinolalia).
IV. Delays in speech development of various origins (due to prematurity, somatic weakness, pedagogical neglect, etc.).
Sensory aphasia(Wernicke's aphasia), or verbal “deafness,” occurs when the left temporal region is damaged (the middle and posterior parts of the superior temporal gyrus). A. R. Luria distinguishes two forms of sensory aphasia: acoustic-gnostic and acoustic-mnestic.
The basis of the defect acoustic-gnostic form constitutes a violation of auditory gnosis. The patient does not differentiate by hearing phonemes that are similar in sound in the absence of deafness (phonemic analysis is considered), as a result of which the understanding of the meaning of individual words and sentences is distorted and impaired. The severity of these disorders may vary. In the most severe cases, the addressed speech is not perceived at all and seems to be speech in a foreign language. This form occurs when the posterior part of the superior temporal gyrus of the left hemisphere is damaged - Brodmann area 22.
In the cerebral cortex, all stimuli that come from the surrounding external and internal environment are analyzed. The largest number of afferent impulses reaches the cells of the 3rd and 4th layers of the cerebral cortex. The cerebral cortex contains centers that regulate the performance of certain functions. I. P. Pavlov considered the cerebral cortex as a set of cortical ends of analyzers. The term “analyzer” refers to a complex complex of anatomical structures, which consists of a peripheral receptor (perceiving) apparatus, conductors of nerve impulses and a center. In the process of evolution, functions are localized in the cerebral cortex. The cortical end of the analyzers is not any strictly defined zone. In the cerebral cortex, a “core” of the sensory system and “scattered elements” are distinguished. The core is the area where the largest number cortical neurons, in which all structures of the peripheral receptor are accurately projected. Scattered elements are located near the core and on different distances From him. If higher analysis and synthesis are carried out in the nucleus, then simpler ones are carried out in scattered elements. At the same time, the zones of “scattered elements” of various analyzers do not have clear boundaries and overlap each other.
Functional characteristics of the cortical zones of the frontal lobe. In the area of the precentral gyrus of the frontal lobe there is the cortical nucleus of the motor analyzer. This area is also called the sensorimotor cortex. Some of the afferent fibers from the thalamus come here, carrying proprioceptive information from the muscles and joints of the body (Fig. 8.7). Descending pathways to the brain stem and spinal cord also begin here, providing the possibility of conscious regulation of movements (pyramidal tracts). Damage to this area of the cortex leads to paralysis of the opposite half of the body.
Rice. 8.7. Somatotopic distribution in the precentral gyrus
The center of writing lies in the posterior third of the middle frontal gyrus. This zone of the cortex gives projections to the nuclei of the oculomotor cranial nerves, and also, through cortico-cortical connections, communicates with the center of vision in the occipital lobe and the control center for the muscles of the arms and neck in the precentral gyrus. Damage to this center leads to impaired writing skills under visual control (agraphia).
The speech motor center (Broca's center) is located in the area of the inferior frontal gyrus. It has pronounced functional asymmetry. When it is destroyed in the right hemisphere, the ability to regulate timbre and intonation is lost, speech becomes monotonous. When the speech motor center on the left is destroyed, speech articulation is irreversibly impaired, up to the loss of the ability to articulate speech (aphasia) and singing (amusia). With partial violations, agrammatism may be observed - the inability to form phrases correctly.
In the region of the anterior and middle third of the upper, middle and partially inferior frontal gyri there is an extensive anterior associative zone of the cortex, which programs complex forms of behavior (planning different forms activities, decision making, analysis of the results obtained, volitional reinforcement of activities, correction of the motivational hierarchy).
The area of the frontal pole and medial frontal gyrus is associated with the regulation of the activity of emotiogenic areas of the brain included in the limbic system, and is related to the control of psycho-emotional states. Disturbances in this area of the brain can lead to changes in what is commonly called “personality structure” and affect a person’s character, his value orientations, and intellectual activity.
The orbital region contains the centers of the olfactory analyzer and is closely connected anatomically and functionally with the limbic system of the brain.
Functional characteristics of the cortical zones of the parietal lobe. In the postcentral gyrus and superior parietal lobule there is the cortical center of the analyzer of general sensitivity (pain, temperature and tactile), or somatosensory cortex. The representation of various parts of the body in it, as in the precentral gyrus, is built according to the somatotopic principle. This principle assumes that body parts are projected onto the surface of the groove in the topographic relationships that they have in the human body. However, the representation of different parts of the body in the cerebral cortex varies significantly. The greatest representation are those areas (hand, head, especially tongue and lips) that are associated with complex movements such as writing, speech, etc. Violations of the cortex in this area lead to partial or complete anesthesia (loss of sensitivity).
Lesions of the cortex in the area of the superior parietal lobule lead to a decrease in pain sensitivity and impairment of stereognosis - recognition of objects by touch without the aid of vision.
In the inferior parietal lobule, in the region of the supramarginal gyrus, there is a center of praxia, which regulates the ability to carry out complexly coordinated actions that form the basis of labor processes, which require special training. A significant number of descending fibers that follow as part of the pathways that control conscious movements (pyramidal pathways) also originate from here. This area of the parietal cortex, through cortico-cortical connections, closely interacts with the frontal cortex and with all sensory areas of the posterior half of the brain.
The visual (optical) speech center is located in the angular gyrus of the parietal lobe. Its damage leads to the inability to understand readable text (alexia).
Functional characteristics of the cortical zones of the occipital lobe. In the area of the calcarine sulcus is the cortical center of the visual analyzer. Its damage leads to blindness. If there are disturbances in the areas of the cortex adjacent to the calcarine sulcus in the region of the occipital pole on the medial and lateral surfaces of the lobe, loss of visual memory, the ability to navigate in an unfamiliar environment may occur, functions associated with binocular vision are disrupted (the ability to use vision to evaluate the shape of objects, the distance to them , correctly proportion movements in space under visual control, etc.).
Functional characteristics of the cortical zones of the temporal lobe. In the area of the superior temporal gyrus, deep in the lateral sulcus, there is the cortical center of the auditory analyzer. Its damage leads to deafness.
The auditory speech center (Wernicke's center) lies in the posterior third of the superior temporal gyrus. Injuries in this area result in an inability to understand oral speech: it is perceived as noise (sensory aphasia).
In the area of the middle and inferior temporal gyri there is a cortical representation of the vestibular analyzer. Damage to this area leads to imbalance when standing and decreased sensitivity of the vestibular apparatus.
Functional characteristics of the cortical zones of the insula.
Information regarding the functions of the insula is contradictory and insufficient. There is evidence that the cortex of the anterior part of the insula is related to the analysis of olfactory and taste sensations, and the posterior part is related to the processing of somatosensory information and auditory perception of speech.
Functional characteristics of the limbic system. Limbic system– a set of a number of brain structures, including the cingulate gyrus, isthmus, dentate and parahippocampal gyri, etc. Participates in the regulation of the functions of internal organs, smell, instinctive behavior, emotions, memory, sleep, wakefulness, etc.
The cingulate and parahippocampal gyri are directly related to the limbic system of the brain (Fig. 8.8 and 8.9). It controls a complex of vegetative and behavioral psycho-emotional reactions to external environmental influences. The cortical representation of the gustatory and olfactory analyzers is located in the parahippocampal gyrus and uncus. At the same time, the hippocampus plays an important role in learning: the mechanisms of short-term and long-term memory are associated with it.

Rice. 8.8. Medial surface of the brain
Basal (subcortical central) nuclei – accumulations of gray matter that form separately lying nuclei that lie closer to the base of the brain. These include the striatum, which constitutes the predominant mass of the hemispheres in lower vertebrates; fence and amygdala (Fig. 8.10).

Rice. 8.9. Limbic system

Rice. 8.10. Basal ganglia
The striatum consists of the caudate and lenticular nuclei. The gray matter of the caudate and lenticular nuclei alternates with layers of white matter, which led to the common name of this group of subcortical nuclei - the striatum.
The caudate nucleus is located lateral and superior to the thalamus, being separated from it by the stria terminalis. The caudate nucleus has a head, body and tail. The lenticular nucleus is located lateral to the caudate. A layer of white matter, the internal capsule, separates the lenticular nucleus from the caudate and from the thalamus. In the lenticular nucleus, the globus pallidus (medially) and the putamen (laterally) are distinguished. The outer capsule (a narrow strip of white matter) separates the shell from the enclosure.
The caudate nucleus, putamen and globus pallidus control complexly coordinated automated movements of the body, control and maintain the tone of skeletal muscles, and are also the highest center for the regulation of such autonomic functions as heat production and carbohydrate metabolism in the muscles of the body. If the putamen and globus pallidus are damaged, slow, stereotypical movements (athetosis) may be observed.
The nuclei of the striatum belong to the extrapyramidal system, which is involved in the control of movements and the regulation of muscle tone.
The fence is a vertical plate of gray matter, the lower part of which continues into the substance of the anterior perforated plate at the base of the brain. The fence is located in the white matter of the hemisphere lateral to the lenticular nucleus and has numerous connections with the cerebral cortex.
The amygdala lies in the white matter of the temporal lobe of the hemisphere, 1.5–2 cm posterior to its temporal pole, through its nuclei it has connections with the cerebral cortex, with the structures of the olfactory system, with the hypothalamus and the nuclei of the brain stem that control the autonomic functions of the body. Its destruction leads to aggressive behavior or an apathetic, lethargic state. Through its connections with the hypothalamus, the amygdala influences the endocrine system as well as reproductive behavior.
The white matter of the hemisphere includes the internal capsule and fibers passing through the cerebral commissures (corpus callosum, anterior commissure, fornix commissure) and heading to the cortex and basal ganglia, fornix, as well as systems of fibers connecting areas of the cortex and subcortical centers within one half of the brain (hemispheres).
I and II lateral ventricles. The cavities of the cerebral hemispheres are the lateral ventricles (I and II), located in the thickness of the white matter under the corpus callosum. Each ventricle consists of four parts: the anterior horn lies in the frontal, the central part - in the parietal, the posterior horn - in the occipital and the inferior horn - in the temporal lobe (Fig. 8.11).
The anterior horns of both ventricles are separated from each other by two plates of a transparent septum. The central part of the lateral ventricle bends from above around the thalamus, forms an arc and passes posteriorly - into the posterior horn, downwards into the inferior horn. The choroid plexus protrudes into the central part and lower horn of the lateral ventricle, which connects to the choroid plexus of the third ventricle through the interventricular foramen.

Rice. 8.11. Ventricles of the brain:
1 - left hemisphere of the brain, 2 - lateral ventricles, 3 - third ventricle, 4 - midbrain aqueduct, 5 - fourth ventricle, 6 - cerebellum, 7 - entrance to the central canal of the spinal cord, 8 - spinal cord
The ventricular system includes paired C-shaped cavities - the lateral ventricles with their anterior, inferior and posterior horns, extending respectively into the frontal lobes, temporal lobes and occipital lobes of the cerebral hemispheres. About 70% of all cerebrospinal fluid is secreted by the choroid plexus of the walls of the lateral ventricles.
From the lateral ventricles, fluid passes through the interventricular foramina into the slit-like cavity of the third ventricle, located in the sagittal plane of the brain and dividing the thalamus and hypothalamus into two symmetrical halves. The cavity of the third ventricle is connected by a narrow canal - the aqueduct of the midbrain (aqueduct of Sylvius) with the cavity of the fourth ventricle. The fourth ventricle communicates through several channels (apertures) with the subarachnoid spaces of the brain and spinal cord.
Diencephalon
The diencephalon is located under the corpus callosum and consists of the thalamus, epithalamus, metathalamus and hypothalamus (Fig. 8.12, see Fig. 7.2).
Thalamus(visual tubercle) – paired, ovoid, formed mainly by gray matter. The thalamus is the subcortical center of all types of sensitivity. The medial surface of the right and left thalami, facing each other, form the lateral walls of the cavity of the diencephalon - the third ventricle; they are connected to each other by an interthalamic fusion. The thalamus contains gray matter, which consists of clusters of neurons that form the thalamic nuclei. The nuclei are separated by thin layers of white matter. About 40 nuclei of the thalamus were studied. The main nuclei are anterior, medial, posterior.

Rice. 8.12. Brain parts
Epithalamus includes the pineal gland, leashes and leash triangles. The pineal body, or pineal gland, which is an endocrine gland, is suspended, as it were, on two leashes, interconnected by a commissure and connected to the thalamus through triangles of leashes. The leash triangles contain nuclei related to the olfactory analyzer. In an adult, the average length of the epiphysis is ~0.64 cm and the mass is ~0.1 g. Metathalamus formed by paired medial and lateral geniculate bodies lying behind each thalamus. The medial geniculate body is located behind the thalamic cushion; it is, along with the lower colliculi of the midbrain roof plate (quadrigeminal), the subcortical center of the auditory analyzer. Lateral - located downward from the pillow, it, together with the upper colliculi of the roof plate, is the subcortical center of the visual analyzer. Cores geniculate bodies connected with the cortical centers of the visual and auditory analyzers.
Hypothalamus, representing the ventral part of the diencephalon, is located anterior to the cerebral peduncles and includes a number of structures that have different origins– the anteriorly located visual part is formed from the telencephalon (optic chiasm, optic tract, gray tubercle, infundibulum, neurohypophysis); from the intermediate - the olfactory part (mammillary bodies and the subthalamic region itself - the hypothalamus) (Fig. 8.13).

Figure 8.13. Basal ganglia and diencephalon
The hypothalamus is the center for the regulation of endocrine functions; it combines nervous and endocrine regulatory mechanisms into a common neuroendocrine system, coordinates nervous and hormonal mechanisms for regulating the functions of internal organs. The hypothalamus contains neurons of the usual type and neurosecretory cells. The hypothalamus and the pituitary gland form a single functional complex, in which the former plays a regulatory and the latter an effector role.
The hypothalamus has more than 30 pairs of nuclei. Large neurosecretory cells of the supraoptic and paraventricular nuclei of the anterior hypothalamic region produce neurosecretes of a peptide nature.
The medial hypothalamus contains neurons that perceive all changes occurring in the blood and cerebrospinal fluid (temperature, composition, hormone content, etc.). The medial hypothalamus is also connected to the lateral hypothalamus. The latter does not have nuclei, but has bilateral connections with the overlying and underlying parts of the brain. The medial hypothalamus is a link between the nervous and endocrine systems. In recent years, enkephalins and endorphins (peptides), which have a morphine-like effect, have been isolated from the hypothalamus. They are believed to be involved in the regulation of behavior and vegetative processes.
Anterior to the posterior perforated substance lie two small spherical mastoid bodies, formed by gray matter covered with a thin layer of white. The nuclei of the mammillary bodies are the subcortical centers of the olfactory analyzer. Anterior to the mastoid bodies is a gray tubercle, which is limited in front by the optic chiasm and the optic tract; it is a thin plate of gray matter at the bottom of the third ventricle, which is extended downward and anteriorly and forms a funnel. The end of it goes into pituitary – an endocrine gland located in the pituitary fossa of the sella turcica. The nuclei of the autonomic nervous system lie in the gray mound. They also influence a person's emotional reactions.
The part of the diencephalon, located below the thalamus and separated from it by the hypothalamic groove, constitutes the hypothalamus itself. The coverings of the cerebral peduncles continue here, the red nuclei and the black substance of the midbrain end here.
III ventricle. Cavity of the diencephalon - III ventricle It is a narrow slit-like space located in the sagittal plane, bounded laterally by the medial surfaces of the thalamus, below by the hypothalamus, in front by the columns of the fornix, the anterior commissure and the terminal plate, behind by the epithalamic (posterior) commissure, above by the fornix, above which the corpus callosum is located. The upper wall itself is formed by the vascular base of the third ventricle, in which its choroid plexus lies.
The cavity of the third ventricle passes posteriorly into the midbrain aqueduct, and in front on the sides through the interventricular foramina communicates with the lateral ventricles.
Midbrain
Midbrain – the smallest part of the brain, lying between the diencephalon and the pons (Fig. 8.14 and 8.15). The area above the aqueduct is called the roof of the midbrain, and on it there are four convexities - the quadrigeminal plate with the superior and inferior colliculi. This is where the visual and auditory reflex pathways go to the spinal cord.
The cerebral peduncles are white round cords that emerge from the pons and move forward to the cerebral hemispheres. The oculomotor nerve (III pair of cranial nerves) emerges from the groove on the medial surface of each peduncle. Each leg consists of a tire and a base, the border between them is a black substance. The color depends on the abundance of melanin in its nerve cells. The substantia nigra belongs to the extrapyramidal system, which is involved in maintaining muscle tone and automatically regulates muscle function. The base of the pedicle is formed by nerve fibers running from the cerebral cortex to the spinal and medulla oblongata and the pons. The tegmentum of the cerebral peduncles contains mainly ascending fibers heading to the thalamus, among which the nuclei lie. The largest are the red nuclei, from which the motor red nucleus-spinal tract begins. In addition, the reticular formation and the nucleus of the dorsal longitudinal fasciculus (intermediate nucleus) are located in the tegmentum.
hindbrain
The hindbrain includes the ventrally located pons and the cerebellum lying behind the pons.

Rice. 8.14. Schematic representation of a longitudinal section of the brain

Rice. 8.15. Transverse section through the midbrain at the level of the superior colliculus (the plane of the section is shown in Fig. 8.14)
Bridge looks like a lying transversely thickened ridge, from the lateral side of which the middle cerebellar peduncles extend to the right and left. The posterior surface of the pons, covered by the cerebellum, participates in the formation of the rhomboid fossa, the anterior surface (adjacent to the base of the skull) borders the medulla oblongata below and the cerebral peduncles above (see Fig. 8.15). It is transversely striated due to the transverse direction of the fibers that go from the pontine nuclei to the middle cerebellar peduncles. On the anterior surface of the bridge along the midline there is a basilar groove located longitudinally, in which the artery of the same name passes.
The bridge consists of many nerve fibers that form pathways, among which are cellular clusters - nuclei. The anterior pathways connect the cerebral cortex with the spinal cord and the cerebellar cortex. In the posterior part of the bridge (tegmentum) there are ascending pathways and partially descending ones, the reticular formation, the nuclei of the V, VI, VII, VIII pairs of cranial nerves are located. On the border between both parts of the bridge lies a trapezoidal body formed by the nuclei and transversely running fibers of the conductive path of the auditory analyzer.
Cerebellum plays a major role in maintaining body balance and coordination of movements. The cerebellum reaches its greatest development in humans in connection with upright posture and the adaptation of the hand to work. In this regard, humans have highly developed hemispheres (new part) of the cerebellum.
In the cerebellum, there are two hemispheres and an unpaired middle phylogenetically old part - the vermis (Fig. 8.16).

Rice. 8.16. Cerebellum: top and bottom views
The surfaces of the hemispheres and the vermis are separated by transverse parallel grooves, between which there are narrow long leaves of the cerebellum. The cerebellum is divided into anterior, posterior and floculonodular lobes, separated by deeper fissures.
The cerebellum consists of gray and white matter. The white matter, penetrating between the gray matter, seems to branch, forming on the median section the figure of a branching tree - the “tree of life” of the cerebellum.
The cerebellar cortex consists of gray matter 1–2.5 mm thick. In addition, in the thickness of the white matter there are accumulations of gray - paired nuclei: dentate nucleus, cork-shaped, spherical and tent nucleus. Afferent and efferent fibers connecting the cerebellum with other parts form three pairs of cerebellar peduncles: the lower ones go to the medulla oblongata, the middle ones to the pons, the upper ones to the quadrigemulus.
By the time of birth, the cerebellum is less developed than the telencephalon (especially the hemisphere), but in the first year of life it develops faster than other parts of the brain. A pronounced enlargement of the cerebellum is observed between the 5th and 11th months of life, when the child learns to sit and walk.
Medulla is a direct continuation of the spinal cord. Its lower boundary is considered to be the place of exit of the roots of the 1st cervical spinal nerve or the decussation of the pyramids, the upper is the posterior edge of the bridge, its length is about 25 mm, its shape approaches a truncated cone, with the base facing upward.
The anterior surface is divided by the anterior median fissure, on the sides of which there are pyramids formed by pyramidal pathways that partially intersect (pyramid decussation) in the depth of the described fissure at the border with the spinal cord. Fibers of the pyramidal tracts connect the cerebral cortex with the nuclei of the cranial nerves and the anterior horns of the spinal cord. On each side of the pyramid there is an olive, separated from the pyramid by the anterior lateral groove.
The posterior surface of the medulla oblongata is divided by the posterior median sulcus; on either side of it there are continuations of the posterior cords of the spinal cord, which diverge upward, passing into the inferior cerebellar peduncles.
The medulla oblongata is built of white and gray matter, the latter is represented by the nuclei of the IX–XII pairs of cranial nerves, olives, centers of respiration and circulation, and the reticular formation. White matter is formed by long and short fibers that make up the corresponding pathways.
Reticular formation is a collection of cells, cell clusters and nerve fibers located in the brain stem (medulla oblongata, pons and midbrain) and forming a network. The reticular formation is connected to all sense organs, motor and sensory areas of the cerebral cortex, the thalamus and hypothalamus, and the spinal cord. It regulates the level of excitability and tone of various parts of the central nervous system, including the cerebral cortex, and is involved in the regulation of the level of consciousness, emotions, sleep and wakefulness, autonomic functions, and purposeful movements.
IV ventricle- This is the cavity of the rhomboid brain; downward it continues into the central canal of the spinal cord. The bottom of the IV ventricle, due to its shape, is called a rhomboid fossa (Fig. 8.17). It is formed by the posterior surfaces of the medulla oblongata and the pons, the upper sides of the fossa are the upper, and the lower are the inferior cerebellar peduncles.

Rice. 8.17. Brainstem; back view. The cerebellum is removed, the rhomboid fossa is open
The median groove divides the bottom of the fossa into two symmetrical halves; on both sides of the groove, medial elevations are visible, expanding in the middle of the fossa into the right and left facial tubercles, where they lie: the nucleus of the VI pair of cranial nerves (abducens nerve), deeper and more lateral – the nucleus of the VII pair ( facial nerve), and downwards the medial eminence passes into the triangle of the hypoglossal nerve, lateral to which is the triangle of the vagus nerve. In the triangles, in the thickness of the brain substance, lie the nuclei of the nerves of the same name. The superior angle of the rhomboid fossa communicates with the midbrain aqueduct. The lateral sections of the rhomboid fossa are called the vestibular fields, where the auditory and vestibular nuclei of the vestibulocochlear nerve (VIII pair of cranial nerves) lie. From the auditory nuclei, transverse medullary stripes extend to the median sulcus, located on the border between the medulla oblongata and the pons and are the fibers of the conductive path of the auditory analyzer. In the thickness of the rhomboid fossa lie the nuclei of the V, VI, VII, VIII, IX, X, XI and XII pairs of cranial nerves.
Blood supply to the brain
Blood enters the brain through two paired arteries: the internal carotid and the vertebral. In the cranial cavity, both vertebral arteries merge, together forming the main (basal) artery. At the base of the brain, the basilar artery merges with the two carotid arteries, forming a single arterial ring (Fig. 8.18). This cascade mechanism of blood supply to the brain ensures sufficient blood flow if any of the arteries fails.

Rice. 8.19. Arteries at the base of the brain and circle of Willis ( right hemisphere cerebellum and right temporal lobe removed); The circle of Willis is shown with a dotted line
Three vessels depart from the arterial ring: the anterior, posterior and middle cerebral arteries, which supply the cerebral hemispheres. These arteries run along the surface of the brain, and from them, blood is delivered deep into the brain by smaller arteries.
The carotid artery system is called the carotid system, which provides 2/3 of the brain's arterial blood needs and supplies the anterior and middle parts of the brain.
The “vertebral-basal” artery system is called the vertebrobasilar system, which provides 1/3 of the needs of the brain and delivers blood to the posterior sections.
The outflow of venous blood occurs mainly through the superficial and deep cerebral veins and venous sinuses (Fig. 8.19). The blood ultimately flows into the internal jugular vein, which exits the skull through the jugular foramen, located at the base of the skull lateral to the foramen magnum.
Meninges
The membranes of the brain protect it from mechanical damage and from the penetration of infections and toxic substances (Fig. 8.20).

Rice. 8.19. Veins and venous sinuses of the brain

Fig.8.20. Coronal section through the skull shell and brain
The first membrane that protects the brain is called the pia mater. It is closely adjacent to the brain, extends into all the grooves and cavities (ventricles) present in the thickness of the brain itself. The ventricles of the brain are filled with a fluid called cerebrospinal fluid or cerebrospinal fluid. The dura mater is directly adjacent to the bones of the skull. Between the soft and hard membranes is the arachnoid (arachnoid) membrane. Between the arachnoid and soft membranes there is a space (subarachnoid or subarachnoid space) filled with cerebrospinal fluid. The arachnoid membrane spreads over the grooves of the brain, forming a bridge, and the soft one merges with them. Due to this, cavities called cisterns are formed between the two shells. The cisterns contain cerebrospinal fluid. These tanks protect the brain from mechanical injuries, acting as “airbags.”
Nerve cells and blood vessels are surrounded by neuroglia - special cellular formations that perform protective, support and metabolic functions, providing the reactive properties of nervous tissue and participating in the formation of scars, inflammatory reactions, etc.
When the brain is damaged, the plasticity mechanism is activated, when the remaining brain structures take over the functions of the affected areas.
Subsequently, the efforts of physiologists were aimed at searching for “critical” areas of the brain, the destruction of which led to disruption of the reflex activity of a particular organ. Gradually, the idea of a rigid anatomical localization of “reflex arcs” emerged, and accordingly the reflex itself began to be thought of as a mechanism of operation of only the lower parts of the brain (spinal centers).
At the same time, the question of the localization of functions in the higher parts of the brain was developed. Ideas about the localization of elements of mental activity in the brain originated a long time ago. In almost every era, certain or
Other hypotheses for the representation of higher mental functions and consciousness in the brain in general.
Austrian physician and anatomist Franz Joseph Gall(1758-1828) amounted to detailed description anatomy and physiology of the human nervous system, equipped with an excellent atlas.
: A whole generation of researchers built on this data. Among Gall's anatomical discoveries are the following: identification of the main differences between the gray and white matter of the brain; determination of the origin of nerves in gray matter; definitive proof of the decussation of the pyramidal tracts and optic nerves; establishment of differences between “convergent” (in modern terminology “associative”) and “divergent” (“projection”) fibers (1808); first clear description of brain commissures; proof of the beginning of the cranial nerves in the medulla oblongata (1808), etc. Gall was one of the first to assign a decisive role to the cerebral cortex in the functional activity of the brain. Thus, he believed that the folding of the brain surface is an excellent solution by nature and evolution to an important problem: ensuring a maximum increase in the surface area of the brain while maintaining its volume more or less constant. Gall introduced the term “arc,” familiar to every physiologist, and described its clear division into three parts.
However, Gall's name is mainly known in connection with his rather dubious (and sometimes scandalous!) doctrine of the localization of higher mental functions in the brain. Attaching great importance to the correspondence between function and structure, Gall back in 1790 made a request to introduce a new science into the arsenal of knowledge - phrenology(from the Greek phren - soul, mind, heart), which also received another name - psychomorphology, or narrow localizationism. As a doctor, Gall observed patients with various disorders of brain activity and noticed that the specifics of the disease largely depended on which part of the brain was damaged. This led him to the idea that each mental function corresponds to a special part of the brain. Seeing the endless variety of characters and individual mental qualities of people, Gall suggested that the strengthening (or greater predominance) in human behavior of any character trait or mental function entails the preferential development of a certain area of the cerebral cortex where this function is represented. Thus, the thesis was put forward: function makes structure. As a result of the growth of this hypertrophied area of the cortex (“brain cone”), pressure on the bones of the skull increases, which, in turn, causes the appearance of an external cranial tubercle above the corresponding area of the brain. In case of underdevelopment of the function, vice versa.
A noticeable depression (“pit”) will appear on the surface of the skull. Using the method of “cranioscopy” created by Gall - studying the relief of the skull using palpation - and detailed “topographical” maps of the surface of the brain, which indicated the locations of all abilities (considered innate), Gall and his followers made a diagnosis, i.e. made a conclusion about character and inclinations of a person, about his mental and moral qualities. Were 2 allocated? areas of the brain where certain abilities of an individual are localized (and 19 of them were recognized as common to humans and animals, and 8 as purely human). In addition to the “bumps” responsible for the implementation of physiological functions, there were also those that testified to visual and auditory memory, orientation in space, a sense of time, and the instinct of procreation; such personal qualities. such as courage, ambition, piety, wit, secrecy, amorousness, caution, self-esteem, sophistication, hope, curiosity, amenability to education, pride, independence, diligence, aggressiveness, fidelity, love of life, love of animals.
Gall's erroneous and pseudoscientific ideas (which were, however, extremely popular in his time) contained a rational grain: recognition of the close connection between the manifestations of mental functions and the activity of the cerebral cortex. The problem of finding differentiated “brain centers” and drawing attention to the functions of the brain was put on the agenda. Gall can truly be considered the founder of “cerebral localization.” Of course, for the further progress of psychophysiology, posing such a problem was more promising than the ancient search for the location of the “common sensory”.
The solution to the question of the localization of functions in the cerebral cortex was facilitated by data accumulating in clinical practice and in animal experiments. German physician, anatomist and physicist Julius Robert Mayer(1814-1878), who practiced for a long time in Parisian clinics, and also served as a ship’s doctor, observed in patients with traumatic brain injuries the dependence of the impairment (or complete loss) of one or another function on damage to a certain area of the brain. This allowed him to suggest that memory is localized in the cerebral cortex (it should be noted that T. Willis came to a similar conclusion back in the 17th century), imagination and judgment are localized in the white matter of the brain, apperception and will are located in the basal ganglia. According to Mayer, a kind of “integral organ” of behavior and psyche is the corpus callosum and the cerebellum.
Over time, clinical study of the consequences of brain damage was supplemented by laboratory artificial extirpation method(from Latin ex(s)tirpatio - removal by root), which allows partially or completely destroying (removing) areas of the brain of animals to determine their functional role in brain activity. At the beginning of the 19th century. They carried out mainly acute experiments on animals (frogs, birds); later, with the development of asepsis methods, they began to carry out chronic experiments, which made it possible to observe the behavior of animals for a more or less long time after surgery. Removal of various parts of the brain (including the cerebral cortex) in mammals (cats, dogs, monkeys) made it possible to elucidate the structural and functional basis of complex behavioral reactions.
It turned out that depriving animals of the higher parts of the brain (birds - the forebrain, mammals - the cerebral cortex) in general did not cause disruption of the basic functions: respiration, digestion, excretion, blood circulation, metabolism and energy. The animals retained the ability to move and react to certain external influences. Consequently, the regulation of these physiological manifestations of vital activity occurs at lower levels (compared to the cerebral cortex) of the brain. However, when the higher parts of the brain were removed, profound changes in the behavior of animals occurred: they became practically blind and deaf, “stupid”; they lost previously acquired skills and could not develop new ones, could not adequately navigate the environment, did not distinguish and could not differentiate objects in the surrounding space. In a word, animals became “living automata” with monotonous and rather primitive ways of responding.
In experiments with partial removal of areas of the cerebral cortex, it was discovered that the brain is functionally heterogeneous and the destruction of one or another area leads to disruption of a certain physiological function. Thus, it turned out that the occipital areas of the cortex are associated with visual function, the temporal areas with auditory function, the area of the sigmoid gyrus with motor function, as well as with skin and muscle sensitivity. Moreover, this differentiation of functions in individual areas of the higher parts of the brain improves as evolutionary development animals.
The strategy of scientific research in the study of brain functions led to the fact that, in addition to the extirpation method, scientists began to use the method of artificial stimulation of certain areas of the brain using electrical stimulation, which also made it possible to assess the functional role of the most important parts of the brain. The data obtained using these laboratory research methods, as well as the results of clinical observations, outlined one of the main directions of psychophysiology in the 19th century. - localization determination nerve centers, responsible for higher mental functions and behavior of the body as a whole. So. in 1861, the French scientist, anthropologist and surgeon Paul Broca (1824-1880), on the basis of clinical facts, decisively spoke out against the physiological equivalence of the cerebral cortex. While autopsying the corpses of patients suffering from a speech disorder in the form of motor aphasia (the patients understood the speech of others, but could not speak themselves), he discovered changes in the posterior part of the inferior (third) frontal gyrus of the left hemisphere or in the white matter under this area of the cortex. Thus, as a result of these observations, Broca established the position of the motor (motor) center of speech, later named after him. In 1874, the German psychiatrist and neurologist K? Wernicke (1848-1905) described the sensory speech center (today bearing his name) in the posterior third of the first temporal gyrus of the left hemisphere. Damage to this center leads to loss of the ability to understand human speech (sensory aphasia). Even earlier, in 1863, using the method of electrical stimulation of certain areas of the cortex (precentral gyrus, precentral region, anterior part of the pericentral lobule, posterior parts of the superior and middle frontal gyri), German researchers Gustav Fritsch and Eduard Hitzig established motor centers (motor cortical fields), irritation of which caused certain contractions of skeletal muscles, "and destruction led to deep disorders of motor behavior. In 4874, the Kiev anatomist and physician Vladimir Alekseevich Betz (1834-1894) discovered efferent nerve cells of motor centers - giant pyramidal cells of layer V cortex, named after him Betz cells. The German researcher Hermann Munch (student of J. Müller and E. Dubois-Reymond) discovered not only motor cortical fields, using the extirpation method he found the centers of sensory perceptions. He was able to show that the center of vision is located in the posterior lobe of the brain, the hearing center is in the temporal lobe. Removal of the occipital lobe of the brain led to the loss of the animal's ability to see (with complete preservation of the visual apparatus). Already in
beginning of the 20th century outstanding Austrian neurologist Constantin Economo(1876-1931) the centers of swallowing and chewing were established in the so-called substantia nigra of the brain (1902), the centers that control sleep were found in the midbrain (1917). Looking ahead a little, let's say that Economo gave an excellent description of the structure of the cerebral cortex an adult and in 1925 refined the cytoarchitectonic map of the cortical fields of the brain, plotting 109 fields on it.
At the same time, it should be noted that in the 19th century. Serious arguments have been put forward against the position of narrow localizationists, according to whose views motor and sensory functions are confined to different areas of the cerebral cortex. Thus, the theory of the equivalence of areas of the cortex arose, affirming the idea of the equal importance of cortical formations for the implementation of any activity of the body - equipotentialism. In this regard, the phrenological views of Gall, one of the most ardent supporters of localizationism, were criticized by the French physiologist Marie Jean Pierre Flourens(1794-1867). Back in 1822, he pointed out the presence of a respiratory center in the medulla oblongata (which he called the “vital node”); connected coordination of movements with the activity of the cerebellum, vision - with the quadrigeminal region; The main function of the spinal cord was to conduct excitation along the nerves. However, despite such seemingly localizationist views, Flourens believed that the basic mental processes (including intellect and will) underlying purposeful human behavior are carried out as a result of the activity of the brain as an integral formation and therefore an integral behavioral function cannot be associated with any particular anatomical formation. Flourens conducted most of his experiments on pigeons and chickens, removing individual parts of their brains and observing changes in the behavior of the birds. Birds' behavior usually recovered some time after surgery, regardless of which areas of the brain were damaged, so Flourens concluded that the degree of impairment of various forms of behavior was determined primarily by how much brain tissue was removed during the operation. Having improved the technique of operations, he was the first to completely remove the forebrain hemispheres of animals and save their lives for further observations.
Based on experiments, Flourens came to the conclusion that the forebrain hemispheres play a decisive role in the implementation of a behavioral act. Their complete removal leads to the loss of all “intelligent” functions. Moreover, particularly severe behavioral disorders were observed in chickens after the destruction of the gray matter on the surface of the cerebral hemispheres - the so-called corticoid plate, an analogue of the mammalian cerebral cortex. Flourens proposed that this area of the brain is the seat of the soul, or "governing spirit", and therefore acts as a single whole, having a homogeneous and equal mass (similar, for example, to the tissue structure of the liver). Despite the somewhat fantastic ideas of equipotentialists, it is worth noting the progressive element in their views. Firstly, complex psychophysiological functions were recognized as the result of the combined activity of brain formations. Secondly, the idea of high dynamic plasticity of the brain, expressed in the interchangeability of its parts, was put forward.
- Gall managed to quite accurately determine the “center of speech,” but it was “officially” discovered by the French researcher Paul Broca (1861).
- In 1842, Mayer, working on determining the mechanical equivalent of heat, came to a generalizing law of conservation of energy.
- Unlike his predecessors, who endowed the nerve with the ability to sense (i.e., recognizing a certain mental quality behind it), Hall considered the nerve ending (in the sensory organ) an “apsychic” formation.
Ideas about the localization of functions in the cerebral cortex are of great practical importance for solving problems of the topic of lesions in the cerebral hemispheres. However, to this day much in this section remains controversial and not entirely resolved. The doctrine of the localization of functions in the cortex has a rather long history - from the denial of the localization of functions in it to the distribution of all functions in the cortex in strictly limited territories human activity, right up to highest qualities the last (memory, will, etc.), and, finally, before returning to the “equipotentiality” of the cortex, i.e. again, in essence, to the denial of the localization of functions (recently abroad).
Ideas about the equivalence (equipotentiality) of various cortical fields conflict with the enormous factual material accumulated by morphologists, physiologists and clinicians. Everyday clinical experience shows that there are certain unshakable natural dependences of functional disorders on the location of the pathological focus. Based on these basic principles, the clinician solves the problems of topical diagnostics. However, this is the case as long as we operate with disorders related to relatively simple functions: movements, sensitivity, etc. In other words, localization in the so-called “projection” zones - cortical fields, directly connected through their paths with the underlying parts of the nervous system and the periphery, is firmly established. The functions of the cortex are more complex, phylogenetically younger, and cannot be narrowly localized; in the implementation complex functions Very large areas of the cortex, and even the entire cortex as a whole, are involved. That is why solving problems of lesion topics based on speech disorders, apraxia, agnosia and, especially, mental disorders, as clinical experience shows, is more difficult and sometimes inaccurate.
At the same time, within the cerebral cortex there are areas whose damage causes one or another character, one or another degree, for example, speech disorders, disorders of gnosis and praxia, the topodiagnostic value of which is also significant. It does not follow from this, however, that there are special, narrowly localized centers that “manage” these most complex forms of human activity. It is necessary to clearly distinguish between the localization of functions and the localization of symptoms.
The foundations of a new and progressive doctrine of the localization of functions in the brain were created by I.P. Pavlov.
Instead of the idea of the cerebral cortex as, to a certain extent, an isolated superstructure above other floors of the nervous system with narrowly localized areas connected along the surface (association) and with the periphery (projection), I.P. Pavlov created the doctrine of the functional unity of neurons belonging to various parts of the nervous system - from receptors in the periphery to the cerebral cortex - the doctrine of analyzers. What we call the center is the highest, cortical, section of the analyzer. Each analyzer is associated with certain areas of the cerebral cortex (Fig. 64).
I.P. Pavlov makes significant adjustments to the previous ideas about the limited territories of cortical centers, to the doctrine of the narrow localization of functions. This is what he says about the projection of receptors into the cerebral cortex.
“Each peripheral receptor apparatus has in the cortex a central, special, isolated territory, as its terminal station, which represents its exact projection. Here, thanks to a special design, perhaps a denser arrangement of cells, more numerous cell connections and the absence of cells of other functions, the most complex irritations occur, are formed (higher synthesis) and their precise differentiation is accomplished (higher analysis). But these receptor elements spread further over a very long distance, perhaps throughout the entire cortex.” This conclusion, based on extensive experimental and physiological studies, is fully consistent with the latest morphological data on the impossibility of accurately delineating cortical cyto-architectonic fields.
Consequently, the functions of the analyzers (or, in other words, the work of the first signaling system) cannot be associated only with the cortical projection zones (nuclei of the analyzers). Moreover, it is impossible to narrowly localize the most complex, purely human functions - the functions of the second signaling system.
I.P. Pavlov defines the functions of human signaling systems as follows. “I imagine the entire complex of higher nervous activity like this. In higher animals, up to and including humans, the first instance for the complex relationships of the organism with the environment is the subcortex closest to the hemispheres with its most complex unconditioned reflexes (our terminology), instincts, drives, affects, emotions (various, usual terminology). These reflexes are caused by relatively few unconditional external agents. Hence the limited orientation in environment and at the same time a weak adaptation.
The second instance is the cerebral hemispheres... Here it arises with the help of a conditional connection (association) new principle activity: signaling of a few, unconditional external agents by a countless mass of other agents, constantly being analyzed and synthesized, making it possible for a very large orientation in the same environment and, at the same time, a much greater adaptation. This constitutes the only signaling system in the animal body and the first in humans.
In a person, another signaling system is added, signaling the first system - speech, its basis or basal component - kinesthetic stimulation of the speech organs. This introduces a new principle of nervous activity - abstraction and together generalization of countless signals from the previous system, in turn, again with the analysis and synthesis of these first generalized signals - a principle that determines limitless orientation in the surrounding world and creates the highest human adaptation - science, both in the form of a universal human empiricism and in its specialized form."
The work of the second signaling system is inextricably linked with the functions of all analyzers, therefore it is impossible to imagine the localization of the complex functions of the second signaling system in any limited cortical fields.
The significance of the legacy left to us by the great physiologist for the correct development of the doctrine of the localization of functions in the cerebral cortex is extremely great. I.P. Pavlov laid the foundations for a new doctrine of the dynamic localization of functions in the cortex. Concepts of dynamic localization suggest the possibility of using the same cortical structures in various combinations to serve various complex cortical functions.
While maintaining a number of clinically established definitions and interpretations, we will try to make some adjustments to our presentation in the light of the teachings of I.P. Pavlova about the nervous system and its pathology.
So, first of all, we need to consider the issue of the so-called projection and association centers. The usual idea of motor, sensory and other projection centers (anterior and posterior central gyri, visual, auditory centers, etc.) is associated with the concept of a rather limited localization in a given area of the cortex of one or another function, and this center is directly related to the underlying neural devices , and subsequently with the periphery, its conductors (hence the definition - “projective”). An example of such a center and its conductor is, for example, the anterior central gyrus and the pyramidal tract; fissura calcarina and radiatio optica, etc. Projection centers are connected through associational paths with other centers, with the surface of the cortex. These broad and powerful association pathways determine the possibility of combined activity of various cortical areas, the establishment of new connections, and, consequently, the formation of conditioned reflexes.
“Association centers,” unlike projection centers, do not have a direct connection with the underlying parts of the nervous system and the periphery; they are connected only with other areas of the cortex, including “projection centers”. An example of an “association center” is the so-called “center of stereognosis” in the parietal lobe, located posterior to the posterior central gyrus (Fig. 65). The posterior central gyrus receives through the thalamo-cortical pathways individual irritations that arise when the hand feels an object: tactile, shape and size (articular-muscular feeling), weight, temperature, etc. All these sensations are transmitted through association fibers from the posterior central gyrus to the “stereognostic center”, where they combine and create a common sensory image of the object. The connections of the “stereognostic center” with the rest of the cortex make it possible to identify and compare this image with the idea of a given object, its properties, purpose, etc., already in memory. (i.e., analysis and synthesis of perception is carried out). This “center,” therefore, does not have a direct connection with the underlying parts of the nervous system and is connected by association fibers with a number of other fields of the cerebral cortex.
The division of centers into projection and association seems to us incorrect. The large hemispheres are a set of analyzers for analyzing, on the one hand, the external world and, on the other, internal processes. The perceptive centers of the cortex seem to be very complicated and geographically extremely widespread. The upper layers of the cerebral cortex, in fact, are entirely occupied by perceptive centers or, in the terminology of I.P. Pavlov, “the brain ends of analyzers.”
From all the lobes, from the lower layers of the cortex, there are efferent conductors that connect the cortical ends of the analyzers with the executive organs through the subcortical, stem and spinal apparatus. An example of such an efferent conductor is the pyramidal tract - this interneuron between the kinesthetic (motor) analyzer and the peripheral motor neuron.
How then, from this point of view, can we reconcile the position about the presence of motor projection centers (in the anterior central gyrus, eye rotation center, etc.), when turned off, a person experiences paralysis, and when irritated, convulsions with a completely clear somatotopic distribution and correspondence? Here we are talking only about damage to the motor projection area for the pyramidal tracts, and not to the “projection motor centers.”
There is no doubt that “voluntary” movements are conditioned motor reflexes, that is, movements that have developed, “trodden down” in the process of individual life experience: but in the development, organization and already created activity of skeletal muscles, everything depends on the afferent device - skin and motor analyzer (clinically - skin and joint-muscular sensitivity, more broadly - kinesthetic sense), without which subtle and precise coordination of a motor act is impossible.
Rice. 64. Cortical sections of analyzers (diagram).
a - outer surface; b - inner surface. Red - skin analyzer; yellow - auditory analyzer: blue - visual analyzer; green - olfactory analyzer; dotted line - motor analyzer.
The motor analyzer (whose task is the analysis and synthesis of “voluntary” movements) does not at all correspond to the idea of cortical motor “projection” centers with definite boundaries of the latter and a clear somatotopic distribution. The motor analyzer, like all analyzers, is connected with very wide areas of the cortex, and the motor function (in relation to “voluntary” movements) is extremely complex (if we take into account not only the determinism of movements and behavior in general, not only the complexity of action complexes, but also the afferent kinesthetic systems , and orientation in relation to the environment and parts of one’s own body in space, etc.).
What does the idea of “projection centers” come down to? It was argued that the latter represented a kind of input or output “trigger gate” for impulses coming into the cortex or emanating from it. And if we accept that “motor projection cortical centers” are only such “gates” (for the broad concept of a motor analyzer is certainly associated with the function of analysis and synthesis), then we should assume that within the anterior central gyrus (and in territories similar to it), and then only in certain of its layers, there is a motor projection area or zone.
How then can we imagine the remaining “projection” centers (skin sensitivity, vision, hearing, taste, smell) associated with other (non-kinesthetic) afferent systems? We think that there is no fundamental difference here: in fact, both in the region of the posterior central gyrus, and within the fissurae calcarinae, etc., impulses flow from the periphery to the cells of a certain layer of the cortex, which is “projected” here, and analysis and synthesis occurs within many strata and wide areas.
Consequently, in each analyzer (its cortical section), including the motor one, there is an area or zone that “projects” to the periphery (motor area) or into which the periphery is “projected” (sensitive areas, including kinesthetic receptors for the motor analyzer ).
It is acceptable that the “projection nucleus of the analyzer” can be identified with the concept of a motor or sensitive projection zone. Maximum violations, wrote I.I. Pavlov, analysis and synthesis arises when precisely such a “projection nucleus” is damaged; If. If we take the maximum dysfunction of the analyzer as a real maximum “breakdown”, which is objectively absolutely correct, then the greatest manifestation of damage to the motor analyzer is central paralysis, and the sensory one is anesthesia. From this point of view, it would be correct to identify the concept of “analyzer core” with the concept of “projection area of the analyzer”.
Rice. 65. Loss of functions observed with damage to various parts of the cerebral cortex (outer surface).
2 - visual disturbances (hemianopsia); 3 - sensitivity disorders; 4 - central paralysis or paresis; 5 - agraphia; 6 - cortical paralysis of gaze and head rotation in the opposite direction; 7 - motor aphasia; 8 - hearing disorders (not observed with unilateral lesions); 9 - amnestic aphasia; 10 - alexia; 11 - visual agnosia (with bilateral damage); 12 - astereognosia; 13 - apraxia; 14 - sensory aphasia.
Based on the above, we consider it correct to replace the concept of a projection center with the concept of a projection area in the analyzer zone. Then the division of cortical “centers” into projection and association is unfounded: there are analyzers (their cortical sections) and, within their limits, projection areas.
Lecture 12. LOCALIZATION OF FUNCTIONS IN THE LARGE HEMISPHERES CORTEX Cortical zones. Projection cortical zones: primary and secondary. Motor (motor) zones of the cerebral cortex. Tertiary cortical zones.
 Loss of functions observed with damage to various parts of the cortex (inner surface). 1 - disorders of smell (not observed with unilateral lesions); 2 - visual disturbances (hemianopsia); 3 - sensitivity disorders; 4 - central paralysis or paresis. Data experimental research by the destruction or removal of certain areas of the cortex and clinical observations indicate that functions are confined to the activity of certain areas of the cortex. An area of the cerebral cortex that has some specific function is called the cortical zone. There are projection, associative cortical zones and motor (motor) zones.
Loss of functions observed with damage to various parts of the cortex (inner surface). 1 - disorders of smell (not observed with unilateral lesions); 2 - visual disturbances (hemianopsia); 3 - sensitivity disorders; 4 - central paralysis or paresis. Data experimental research by the destruction or removal of certain areas of the cortex and clinical observations indicate that functions are confined to the activity of certain areas of the cortex. An area of the cerebral cortex that has some specific function is called the cortical zone. There are projection, associative cortical zones and motor (motor) zones.
 The projection cortical zone is the cortical representation of the analyzer. Neurons of projection zones receive signals of one modality (visual, auditory, etc.). There are: - primary projection zones; - secondary projection zones, providing an integrative function of perception. In the zone of a particular analyzer, tertiary fields, or associative zones, are also distinguished.
The projection cortical zone is the cortical representation of the analyzer. Neurons of projection zones receive signals of one modality (visual, auditory, etc.). There are: - primary projection zones; - secondary projection zones, providing an integrative function of perception. In the zone of a particular analyzer, tertiary fields, or associative zones, are also distinguished.
 The primary projection fields of the cortex receive information mediated through the smallest number of switches in the subcortex (thalamus, diencephalon). The surface of peripheral receptors is, as it were, projected onto these fields. Nerve fibers enter the cerebral cortex mainly from the thalamus (these are afferent inputs).
The primary projection fields of the cortex receive information mediated through the smallest number of switches in the subcortex (thalamus, diencephalon). The surface of peripheral receptors is, as it were, projected onto these fields. Nerve fibers enter the cerebral cortex mainly from the thalamus (these are afferent inputs).
 The projection zones of the analyzing systems occupy the outer surface of the posterior cortex of the brain. This includes the visual (occipital), auditory (temporal) and sensory (parietal) areas of the cortex. The cortical department also includes the representation of taste, olfactory, visceral sensitivity
The projection zones of the analyzing systems occupy the outer surface of the posterior cortex of the brain. This includes the visual (occipital), auditory (temporal) and sensory (parietal) areas of the cortex. The cortical department also includes the representation of taste, olfactory, visceral sensitivity
 Primary sensory areas (Brodmann areas): visual - 17, auditory - 41 and somatosensory - 1, 2, 3 (collectively they are called sensory cortex), motor (4) and premotor (6) cortex
Primary sensory areas (Brodmann areas): visual - 17, auditory - 41 and somatosensory - 1, 2, 3 (collectively they are called sensory cortex), motor (4) and premotor (6) cortex
 Primary sensory areas (Brodmann areas): visual - 17, auditory - 41 and somatosensory - 1, 2, 3 (collectively they are called sensory cortex), motor (4) and premotor (6) cortex Each field of the cerebral cortex is characterized by a special composition neurons, their location and connections between them. The fields of the sensory cortex, in which the primary processing of information from sensory organs occurs, differ sharply from the primary motor cortex, which is responsible for generating commands for voluntary muscle movements.
Primary sensory areas (Brodmann areas): visual - 17, auditory - 41 and somatosensory - 1, 2, 3 (collectively they are called sensory cortex), motor (4) and premotor (6) cortex Each field of the cerebral cortex is characterized by a special composition neurons, their location and connections between them. The fields of the sensory cortex, in which the primary processing of information from sensory organs occurs, differ sharply from the primary motor cortex, which is responsible for generating commands for voluntary muscle movements.
 In the motor cortex, neurons shaped like pyramids predominate, and the sensory cortex is represented mainly by neurons whose body shape resembles grains or granules, which is why they are called granular. Structure of the cerebral cortex I. molecular II. external granular III. external pyramidal IV. internal granular V. ganglionic (giant pyramids) VI. polymorphic
In the motor cortex, neurons shaped like pyramids predominate, and the sensory cortex is represented mainly by neurons whose body shape resembles grains or granules, which is why they are called granular. Structure of the cerebral cortex I. molecular II. external granular III. external pyramidal IV. internal granular V. ganglionic (giant pyramids) VI. polymorphic
 Neurons of the primary projection zones of the cortex generally have the highest specificity. For example, neurons in the visual areas selectively respond to shades of color, direction of movement, character of lines, etc. However, in the primary zones of individual areas of the cortex there are also multimodal type neurons that respond to several types of stimuli and neurons whose reaction reflects the influence of nonspecific ( limbicoreticular) systems.
Neurons of the primary projection zones of the cortex generally have the highest specificity. For example, neurons in the visual areas selectively respond to shades of color, direction of movement, character of lines, etc. However, in the primary zones of individual areas of the cortex there are also multimodal type neurons that respond to several types of stimuli and neurons whose reaction reflects the influence of nonspecific ( limbicoreticular) systems.
 Projection afferent fibers end in the primary fields. Thus, fields 1 and 3, occupying the medial and lateral surfaces of the posterior central gyrus, are the primary projection fields of cutaneous sensitivity of the body surface.
Projection afferent fibers end in the primary fields. Thus, fields 1 and 3, occupying the medial and lateral surfaces of the posterior central gyrus, are the primary projection fields of cutaneous sensitivity of the body surface.
 The functional organization of projection zones in the cortex is based on the principle of topical localization. Perceptive elements located next to each other in the periphery (for example, areas of the skin) are projected onto the cortical surface also next to each other.
The functional organization of projection zones in the cortex is based on the principle of topical localization. Perceptive elements located next to each other in the periphery (for example, areas of the skin) are projected onto the cortical surface also next to each other.
 The lower limbs are represented in the medial part, and projections of the receptor fields of the skin surface of the head are located lowest on the lateral part of the gyrus. In this case, areas of the body surface richly equipped with receptors (fingers, lips, tongue) are projected onto large area cortex than areas with fewer receptors (thigh, back, shoulder).
The lower limbs are represented in the medial part, and projections of the receptor fields of the skin surface of the head are located lowest on the lateral part of the gyrus. In this case, areas of the body surface richly equipped with receptors (fingers, lips, tongue) are projected onto large area cortex than areas with fewer receptors (thigh, back, shoulder).

 Fields 17-19, located in the occipital lobe, are the visual center of the cortex; field 17, occupying the occipital pole itself, is primary. The 18th and 19th fields adjacent to it perform the function of secondary fields and receive inputs from the 17th field.
Fields 17-19, located in the occipital lobe, are the visual center of the cortex; field 17, occupying the occipital pole itself, is primary. The 18th and 19th fields adjacent to it perform the function of secondary fields and receive inputs from the 17th field.
 The auditory projection fields are located in the temporal lobes (41, 42). Next to them, on the border of the temporal, occipital and parietal lobes, are located the 37th, 39th and 40th, characteristic only of the human cerebral cortex. For most people, these fields of the left hemisphere contain the speech center, which is responsible for the perception of oral and written speech.
The auditory projection fields are located in the temporal lobes (41, 42). Next to them, on the border of the temporal, occipital and parietal lobes, are located the 37th, 39th and 40th, characteristic only of the human cerebral cortex. For most people, these fields of the left hemisphere contain the speech center, which is responsible for the perception of oral and written speech.

 Secondary projection fields, receiving information from the primary ones, are located next to them. The neurons of these fields are characterized by the perception of complex signs of stimuli, but at the same time the specificity corresponding to the neurons of the primary zones is preserved. The complication of the detector properties of neurons in the secondary zones can occur through the convergence of neurons in the primary zones on them. In the secondary zones (18th and 19th Brodmann fields) detectors appear more complex elements contour: edges of limited line lengths, corners with different orientations, etc.
Secondary projection fields, receiving information from the primary ones, are located next to them. The neurons of these fields are characterized by the perception of complex signs of stimuli, but at the same time the specificity corresponding to the neurons of the primary zones is preserved. The complication of the detector properties of neurons in the secondary zones can occur through the convergence of neurons in the primary zones on them. In the secondary zones (18th and 19th Brodmann fields) detectors appear more complex elements contour: edges of limited line lengths, corners with different orientations, etc.
 Motor (motor) zones of the cerebral cortex are areas of the motor cortex, the neurons of which cause a motor act. The motor areas of the cortex are located in the precentral gyrus of the frontal lobe (in front of the projection zones of cutaneous sensitivity). This part of the cortex is occupied by fields 4 and 6. From the V layer of these fields, the pyramidal tract originates, ending on the motor neurons of the spinal cord.
Motor (motor) zones of the cerebral cortex are areas of the motor cortex, the neurons of which cause a motor act. The motor areas of the cortex are located in the precentral gyrus of the frontal lobe (in front of the projection zones of cutaneous sensitivity). This part of the cortex is occupied by fields 4 and 6. From the V layer of these fields, the pyramidal tract originates, ending on the motor neurons of the spinal cord.
 Premotor zone (field 6) The premotor zone of the cortex is located in front of the motor zone, it is responsible for muscle tone and coordinated movements of the head and torso. The main efferent outputs from the cortex are the axons of layer V pyramids. These are efferent, motor neurons involved in the regulation of motor functions.
Premotor zone (field 6) The premotor zone of the cortex is located in front of the motor zone, it is responsible for muscle tone and coordinated movements of the head and torso. The main efferent outputs from the cortex are the axons of layer V pyramids. These are efferent, motor neurons involved in the regulation of motor functions.


 Tertiary or interanalyzer zones (associative) Prefrontal zone (fields 9, 10, 45, 46, 47, 11), parietotemporal (fields 39, 40) Afferent and efferent projection zones of the cortex occupy a relatively small area. Most of the surface of the cortex is occupied by tertiary or interanalyzer zones, called associative zones. They receive multimodal inputs from the sensory areas of the cortex and thalamic associative nuclei and have outputs to the motor areas of the cortex. Associative zones provide integration of sensory inputs and play a significant role in mental activity (learning, thinking).
Tertiary or interanalyzer zones (associative) Prefrontal zone (fields 9, 10, 45, 46, 47, 11), parietotemporal (fields 39, 40) Afferent and efferent projection zones of the cortex occupy a relatively small area. Most of the surface of the cortex is occupied by tertiary or interanalyzer zones, called associative zones. They receive multimodal inputs from the sensory areas of the cortex and thalamic associative nuclei and have outputs to the motor areas of the cortex. Associative zones provide integration of sensory inputs and play a significant role in mental activity (learning, thinking).
 Functions of various areas of the neocortex: 5 3 7 6 4 1 2 Memory, needs Triggering behavior 1. Occipital lobe - visual cortex. 2. Temporal lobe – auditory cortex. 3. Anterior part of the parietal lobe – pain, skin and muscle sensitivity. 4. Inside the lateral sulcus (insula) – vestibular sensitivity and taste. 5. The posterior part of the frontal lobe is the motor cortex. 6. The posterior part of the parietal and temporal lobes is the associative parietal cortex: it combines signal flows from different sensory systems, speech centers, and thinking centers. 7. Anterior part of the frontal lobe - associative frontal cortex: taking into account sensory signals, signals from the centers of needs, memory and thinking, makes decisions on the launch of behavioral programs (“center of will and initiative”).
Functions of various areas of the neocortex: 5 3 7 6 4 1 2 Memory, needs Triggering behavior 1. Occipital lobe - visual cortex. 2. Temporal lobe – auditory cortex. 3. Anterior part of the parietal lobe – pain, skin and muscle sensitivity. 4. Inside the lateral sulcus (insula) – vestibular sensitivity and taste. 5. The posterior part of the frontal lobe is the motor cortex. 6. The posterior part of the parietal and temporal lobes is the associative parietal cortex: it combines signal flows from different sensory systems, speech centers, and thinking centers. 7. Anterior part of the frontal lobe - associative frontal cortex: taking into account sensory signals, signals from the centers of needs, memory and thinking, makes decisions on the launch of behavioral programs (“center of will and initiative”).
 Individual large association areas are located next to the corresponding sensory areas. Some associative zones perform only a limited specialized function and are connected to other associative centers capable of subjecting information to further processing. For example, the auditory association area analyzes sounds, categorizing them, and then transmits signals to more specialized areas, such as the speech association area, where the meaning of words heard is perceived.
Individual large association areas are located next to the corresponding sensory areas. Some associative zones perform only a limited specialized function and are connected to other associative centers capable of subjecting information to further processing. For example, the auditory association area analyzes sounds, categorizing them, and then transmits signals to more specialized areas, such as the speech association area, where the meaning of words heard is perceived.
 The association fields of the parietal lobe combine information coming from the somatosensory cortex (from the skin, muscles, tendons and joints regarding body position and movement) with visual and auditory information coming from the visual and auditory cortices of the occipital and temporal lobes. This combined information helps you have an accurate understanding of your own body while moving around in the environment.
The association fields of the parietal lobe combine information coming from the somatosensory cortex (from the skin, muscles, tendons and joints regarding body position and movement) with visual and auditory information coming from the visual and auditory cortices of the occipital and temporal lobes. This combined information helps you have an accurate understanding of your own body while moving around in the environment.
 Wernicke's area and Broca's area are two areas of the brain involved in the process of reproducing and understanding information related to speech. Both areas are located along the Sylvian fissure (the lateral fissure of the cerebral hemispheres). Aphasia is a complete or partial loss of speech caused by local lesions of the brain.
Wernicke's area and Broca's area are two areas of the brain involved in the process of reproducing and understanding information related to speech. Both areas are located along the Sylvian fissure (the lateral fissure of the cerebral hemispheres). Aphasia is a complete or partial loss of speech caused by local lesions of the brain.






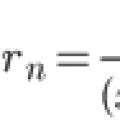 Taylor series expansion Approximate solution of the Cauchy problem for the ordinary
Taylor series expansion Approximate solution of the Cauchy problem for the ordinary What is NOT taught at school What is not taught at school ask
What is NOT taught at school What is not taught at school ask Money Thinking Formula (A
Money Thinking Formula (A